Fossilised Biomolecules and Biomarkers in Carbonate Concretions from Konservat-Lagerstätten


1
Western Australia Organic and Isotope Geochemistry Centre, The Institute for Geoscience Research, School of Earth and Planetary Sciences, Curtin University, GPO Box U1987, Perth, WA 6845, Australia
2
The Commonwealth Scientific and Industrial Research Organisation (CSIRO), Mineral Resources, Kensington, Perth, WA 6151, Australia
*
Author to whom correspondence should be addressed.
Minerals 2019, 9(3), 158; https://doi.org/10.3390/min9030158
Submission received: 20 January 2019
/
Revised: 24 February 2019
/
Accepted: 27 February 2019
/
Published: 6 March 2019
(This article belongs to the Special Issue Microbialites: Preservation of Extant and Extinct Systems)
Abstract
:In the vast majority of fossils, the organic matter is degraded with only an impression or cast of the organism remaining. In rare cases, ideal burial conditions result in a rapid fossilisation with an exceptional preservation of soft tissues and occasionally organic matter. Such deposits are known as Lagerstätten and have been found throughout the geological record. Exceptional preservation is often associated with finely crystalline quartz (e.g., cherts), fine sediments (e.g., muds) or volcanic ashes. Other mechanisms include burial in anoxic/euxinic sediments and in the absence of turbidity or scavenging. Exceptional preservation can also occur when an organism is encapsulated in carbonate cement, forming a concretion. This mechanism involves complex microbial processes, resulting in a supersaturation in carbonate, with microbial sulfate reduction and methane cycling the most commonly suggested processes. In addition, conditions of photic zone euxinia are often found to occur during concretion formation in marine environments. Concretions are ideal for the study of ancient and long-extinct organisms, through both imaging techniques and biomolecular approaches. These studies have provided valuable insights into the evolution of organisms and their environments through the Phanerozoic and have contributed to increasing interest in fields including chemotaxonomy, palaeobiology, palaeoecology and palaeophysiology.
Keywords:
concretions; preservation; biomarkers; soft tissue; evolution; chlorobi; lagerstätten; fossilisation1. Introduction
Fossils play a significant role in defining the geologic timescale which provides a framework for understanding important events in Earth’s history and the evolution of organisms. They also provide information on past climates and environments and, in some cases, on past ecosystems and evolution. Before fossilisation, organic biological remains (e.g., soft tissues) are usually degraded and rarely survive to provide any useful information [1]. In some exceptionally preserved fossils, the degradation of organic matter appears to have stalled early in the fossilisation process. Protection against the inorganic environment through isolation and cementation is thought to have prevented the complete mineralisation of soft tissue (i.e., authigenic mineralization) and molecules (i.e., degradation to CO2 and CH4), allowing them to be preserved for tens of millions of years (Ma) [2]. A better understanding of these exceptional preservation processes will allow the targeting of favourable specimens, providing much more information to address fundamental questions related to palaeoenvironments, evolution and the extinction of life [3].
For example, the unique burial conditions in an anaerobic lake paved the way for the preservation of hemoglobin-derived compounds that were found inside a fossil mosquito (Middle Eocene, 46 Ma) and provided evidence of its hematophagic lifestyle [4]. More recently, red blood cells, along with amino acids from collagen, were reported in 75-Ma-old dinosaur vertebrate bones [5]. Such observations reveal biological features that could offer insights into relationships, physiology and behaviour of long-extinct animals. This level of preservation was proven to be possible over much longer geological timescales with the in situ detection of collagen in a 195-Ma-old dinosaur bone [6] and the release of preserved soft tissues from the decalcification of animal bones up to 205 Ma [7]. However, the majority of studies showing an exceptional preservation of soft tissue, cellular remains and/or biomolecules were performed on bones of extinct vertebrates and, therefore, provide only a limited view into palaeoecology, palaeobiodiversity, palaeophysiology and complex palaeodepositional conditions.
In contrast, recent pioneering studies have shown that a much wider biological range of exceptionally preserved soft tissue, cellular remains and molecules are encapsulated inside the so-called carbonate concretions that are found in sediments of all geological ages [8,9,10]. Notably, the oldest-known intact lipids (cholesterol) were reported inside a 380-Ma-old Devonian concretion [10]. Moreover, the detection of cholesterol, red and white blood cells and collagen in a 183-Ma-old fossil vertebra bone encapsulated in a carbonate concretion provided unprecedented insights into the predatory lifestyle and diet of an ichthyosaur-like reptile [8].
In this review, we summarise the mechanisms leading to the exceptional preservation of fossils with a special focus on carbonate concretions. We give an overview of the formation of concretions and the role of microorganisms and review the analytical techniques that have allowed palaeobiological and palaeoecological information to be obtained from fossils in concretions. For a more extensive discussion of exceptionally preserved fossils in general, the reader is directed to reviews by Cohen and Macdonald [11] and Muscente et al. [12].
2. Modes of Preservation
In the majority of fossils, complex biogeochemical and geological processes result in the destruction of almost all organic matter present in a deceased organism. These processes commence immediately after the death of the organism and include degradation within the water column (e.g., scavenging) as well as sedimentary processes. Immediately after deposition but before burial, microbial activity begins to degrade organic matter. Over longer time scales, geological processes contribute to the destruction of organic matter during the burial and exhumation of a sediment. Mechanisms such as authigenic mineralisation and permineralisation, the maturation of organic matter during diagenesis and metagenesis, and finally exhumation and weathering strongly affect the preservation of organic matter [1]. However in some exceptional cases, the preservation of organic matter may occur through the rapid precipitation and neoformation of minerals from saturated ionic solutions [13]. In such cases microbial activity often plays a pivotal role, establishing steep geochemical gradients around the organism being fossilised [13,14]. Delicate structural information may be preserved as the carbonaceous residue of resistant tissues, such as cell walls and cuticles, as seen in remnants of land plants in the Rhynie cherts of Scotland [15] or in silicified tree trunks in the Petrified Forest National Park, USA [16], but the original tissue is most often fully replaced.
“Exceptional” fossil preservation occurs only in rare depositional conditions. The mechanisms of exceptional preservation can include entombment or primary cementation in concretions of calcite, siderite or dolomite; in fossil resins (fossilised plant exudates); or as carbonaceous impressions, preserving the carbon residue of the decayed organism. Some specific circumstances, including rapid burial, slow decaying processes, anoxic/euxinic conditions and the rapid microbial-mediated entombment of decaying organisms, lead to the preservation of biomolecules [13]. Such circumstances favour the exceptional preservation of biomolecules and molecular fossils [9]. It has also been shown that a mechanism of preservation involving microbial mats occurred in the Mesoproterozoic through Ediacaran [17].
The two types of exceptionally preserved fossil deposits found in the rock record are known as Konservat-Lagerstätten and Konzentrat-Lagerstätten [18]. Konservat-Lagerstätten (conservation Lagerstätten) refer to “exceptional” fossil deposits which preserve fossils and soft-bodied organisms [10,19], providing important insights into significant time periods in the evolution and history of life. These deposits are formed by rapid burial and early diagenetic cementation, occurring under anoxic to euxinic conditions—processes which prevent the decay of both soft and hard tissue [19]. The study of these fossils can yield important information on the evolution of species and past environmental conditions [20]. The unique taphonomy of fossils is site-dependent. A well-known example of a Konservat-Lagerstätten is the Cambrian Burgess Shale (British Columbia, Canada), which was discovered in 1909 by the palaeontologist Charles Walcott [21]. This site is renowned for its high diversity of exceptionally preserved soft and hard fossilised organisms, preserving an assemblage of some of the earliest animal species, and is closely associated with the Cambrian explosion and development of multicellular life on Earth [21]. Conversely, Konzentrat-Lagerstätten (concentration Lagerstätten) are concentrated fossil deposits that do not tend to exceptionally preserve the soft tissues of organisms, containing an accumulation of organic hard parts such as bone beds, with some exceptions, for example, reef beds which are formed in situ [2,20,22]. Notable examples of Konservat-Lagerstätten and Konzentrat-Lagerstätten throughout the geological record are summarised below (Table 1). See Muscente et al. [12] for a more complete list.
“Selective” preservation (decay resistance) can also occur, for example, for a few high molecular-weight organic materials [48]. Some specific resistant biopolymeric constituents have been found in higher plant leaf cuticles [49] as well as in the cell walls of marine and freshwater algae [50,51,52,53].
3. Molecules of Life in Concretions
One mechanism of exceptional preservation is encasement within carbonate concretions. These are mineral structures formed by the cementation of carbonate around a nucleus, which can often be organic material in the form of soft tissue [54]. Carbonate concretions have been found to preserve the soft tissues of plants and animals throughout sedimentary rocks of all geological ages [54,55]. Carbonate concretions have been extensively studied [8,9,10,35,56], as they have the potential to contain an abundance of information regarding the conditions allowing for the exceptional preservation of fossils [57] (Figure 1).
The exceptional preservation of biological remains inside concretions is attributed to carbonate precipitation induced by microbial activity surrounding a decaying carcass or plant. This microbially induced carbonate deposition, commencing immediately upon decay, prevents the full decomposition of the enclosed biological material for tens to hundreds of millions of years [8]. In nature, many microorganisms are known to induce carbonate precipitation by altering the solution chemistry through a wide range of physiological activities such as photosynthesis, ureolysis, ammonification, denitrification, sulfate reduction, anaerobic sulfide oxidation and methane oxidation [58]. Microbially formed biofilms are believed to surround the decaying biomass and to serve as a template for carbonate concretion growth. These biofilms consist of extracellular polymeric substances (EPS) comprised of sugars, proteins, DNA and large amounts of trapped divalent cations such as Ca2+ and act as effective nucleation sites for carbonate precipitation [14,59]. Recent biomarker (molecular fossil) studies on carbonate concretions have provided initial insights into the palaeodepositional environments that led to the encapsulation and preservation of organic matter as well as information on the identity and function of microorganisms involved in the process [10,60]. These studies provided evidence for the presence of anoxic sulfidic conditions and an active microbial sulfur cycle at the time of organic matter preservation. Melendez et al. [10] unequivocally established that anoxygenic photosynthesis performed by green sulfur bacteria using hydrogen sulfide as an electron donor played a pivotal role in the preservation of biomolecules 380-Ma-years ago. Microbial sulfate reduction in the sediments and overlying bottom waters was ascribed as the source of this hydrogen sulfide and provided persistent anaerobic conditions allowing for the excellent preservation of lipids in Phanerozoic environments (541 Ma to present). While these lipid biomarker studies provided evidence that highly sulfidic conditions contributed to the preservation of organic matter in carbonate concretions, the microbes and processes that resulted in the actual carbonate deposition surrounding the fossil organic matter remains to be fully understood.
A combination of many processes and factors leads to the formation of carbonate concretions [33,61]. The growth of concretions can occur by the precipitation of carbonate cement [54] via three different mechanisms: pervasive growth, concentric growth from inside to outside [62] or concentric growth from outside to inside [61]. The growth of carbonate concretions has also been shown to be influenced by microbial processes [61,63], including Fe (III) reduction, sulfate reduction and methanogenesis [61]. These microbial processes result in carbonate supersaturation as they yield CO2 or dissolved CO32−, as well as influence the composition of the pore water [61]. In marine environments, sulfate reduction occurs preferentially over methanogenesis, due to high amounts of Mg2+ and sulfate-reducing bacteria [64]. This sulfate reduction results in the consumption of dissolved iron in porewaters, preferentially forming pyrite [55,65] over iron carbonate; hence, calcium carbonates are most commonly found in marine environments. Information on the conditions and locations of carbonate concretion formation can be determined from the amounts of pyrite, carbon and sulfur in concretions [65], as the amount of sulfide present is related to the timing of pyrite formation relative to concretion formation [66]. Conversely, in brackish to freshwater conditions, methanogenesis is supported by anaerobic sediments due to a low amount of sulfate reduction [55,64]; the reaction that occurs between iron compounds and organic matter results in the formation of iron carbonate concretions [31,67,68,69]. Iron carbonate is preferentially formed over calcium carbonate as the decreased amount of sulfate reduction occurring means that the reaction of iron with sulfate to form pyrite is minimal [55].
Traces of ancient life can be preserved in sedimentary settings either as morphological fossils or molecular fossils (lipid biomarkers, pigments, porphyrins, proteins or DNA) [70]. These molecules are stable and often well-preserved in sediments where hard fossil parts are absent, making them highly valuable markers of past life. Biomarkers are related to the lipids and other biochemicals of extant biological organisms, thus allowing their biomarker–precursor relationships to be established from the three domains life: eukaryotes, bacteria and archaea [70,71,72]. The identity, isomeric arrangement and stable isotopic composition of molecular biomarkers are widely used to provide palaeoenvironmental insights into deposition [71]. With the advance of analytical technologies, including novel methodologies for sequencing ancient DNA and proteins [73,74,75], such studies are providing an increasingly greater resolution. As analytical methods have progressed over the last decade, so has our knowledge of the chemical pathways involved in the formation of biomarkers and the specific palaeoenvironmental conditions required for their effective preservation [71].
4. Imaging Techniques
4.1. Nondestructive Tomographic Methods
Although three-dimensional tomography techniques started to evolve during the beginning of the twentieth century, this has historically been a slow and cumbersome process [76]. In recent years, however, the development of high resolution and rapid technologies promoted the development of the new field of “virtual palaeontology”. Initially developed for medical research, these technologies have been progressively applied to investigate palaeontological samples. Magnetic resonance imaging, for instance, can reveal anatomical details at the scale of a dozen micrometres and can provide insights into the presence, or the absence, of organic residues [77]. In addition, computed X-ray microtomography has been particularly important in contributing to the taxonomic classification of fossil bones [78]. Although nondestructive tomographic methods have only been employed in rare occasions to investigate fossils preserved within carbonate concretions or nodules [79], synchrotron-based tomography holds great potential to image fossils encapsulated within concretions [80,81,82]. An exhaustive description of these nondestructive tomographic techniques can be found in References [76,83,84]. However, when the host concretion and the fossil preserved within share a similar mineralogy, destructive techniques, coupled with high resolution imaging and software reconstruction, remain the most efficient for fossil characterisation [85].
4.2. Microbeam X-ray Fluorescence (XRF) Mapping
Benchtop microbeam XRF mapping is applicable to flat surfaces (e.g., polished samples/ thin sections), allowing for a rapid determination of the elemental distribution. The spot size of 25 µm allows for a comprehensive overview of elemental distributions in the sample. This technique reveals, within hours and with minimal processing time, complex elemental distributions that can be used to infer the mineralogical composition of concretion matrix and fossils [8]. This rapid screening method is particularly beneficial to determine areas of interest which should be further characterised at higher resolution. Such additional, highly detailed methods include, for instance, high resolution synchrotron-based XRF microprobe (SRXRF) providing elemental mapping with a 2 µm beam-spot size [86].
4.3. Optical and Scanning Electron Microscopy (SEM)
Carbonate concretions are commonly investigated using traditional optical microscopy. Performed on thin sections, optical microscopy is particularly useful to investigate the texture of minerals forming the concretions and to provide insight into the mode of concretion formation [87,88,89].
Scanning electron microscopy (SEM) coupled with energy dispersion spectrometry (EDS) are particularly useful to further characterise the elemental distribution and mineralogy of microstructures. SEM can also be coupled with other techniques, for example, EDS and Time of Flight Secondary Ion Mass Spectrometry (ToF-SIMS) for chemical information and Focused Ion Beam (FIB) for the spatial relationship between elements of interest.
ToF-SIMS uses a high-energy beam of ions (primary ions) to break down the surface layers of a sample into secondary ions which are analysed by mass spectrometry, producing spatially resolved chemical maps of both organic and inorganic samples [90,91]. This technique has been applied to the analysis of organics in well-preserved fossils, including hemoglobin-derived porphyrins in a Middle Eocene mosquito [4], aliphatic and aromatic carbons in the vascular structures of early land plants in the Devonian Rhynie chert [92], collagen in a Campanian hadrosaur bone [93] and fatty acyl biomarkers in a Late Oligocene whale bone [94]. ToF-SIMS has also been applied to distinguish well-preserved melanosomes (melanin-containing organelles) from potential fossilised bacteria [95,96] and to determine the distribution of 12C and 13C associated with red blood cell-like structures in a vertebra fossil encapsulated in a carbonate concretion [8]. FIB-SEM also presents the potential to be used to prepare delicate samples for further analyses using transmission electron microscopy (TEM) which provides additional chemical and textural information at a nanometer scale. TEM is a common technique in the nanoscale investigation of microstructures of proposed biological origin [97].
These imaging techniques give insights into the geochemistry of the specimen at a very high resolution. They complement other traditional and destructive geochemical approaches used in organic geochemistry, as illustrated in Figure 2. In a standard organic geochemistry workflow, the fossil sample is ground to a fine powder and the organic molecules are isolated by extracting with a suitable mixture of solvents [98]. The extractable organics are typically separated on the basis of polarity using silica gel column chromatography, and the saturated, aromatic and polar fractions analysed using techniques such as gas chromatography-mass spectrometry (GC-MS) and gas chromatograph-isotope ratio mass spectrometry (GC-irMS), e.g., in References [35,99]. The polar organic fractions may be desulfurised by a treatment with Raney nickel, which breaks carbon–sulfur bonds, releasing organic matter that was sulfurised during early diagenesis [10].
5. The Role of Biomarkers in Concretions
Biomarkers are molecular fossils formed by the transformation of hydrocarbon lipids in sediments to hydrocarbon skeletons through the processes of eogenesis and diagenesis [98]. They are significant in that they are able to provide biological information from organisms which lived hundreds of millions of years ago, as they retain the structure of the original lipids from which they are derived [98]. As such, biomarkers allow for the detailed investigation of conditions and processes which allow for exceptional fossil preservation [9,55], as well as provide important information regarding microbial systems and preservation conditions [10,35,60]. Similar to biomarkers, DNA can provide a breadth of information on fossilised species and preservation conditions; however, it is limited in its usefulness as it can only survive for up to one million years in sediments [100], whereas biomarkers have been shown to survive for over one billion years in ideal conditions [101].
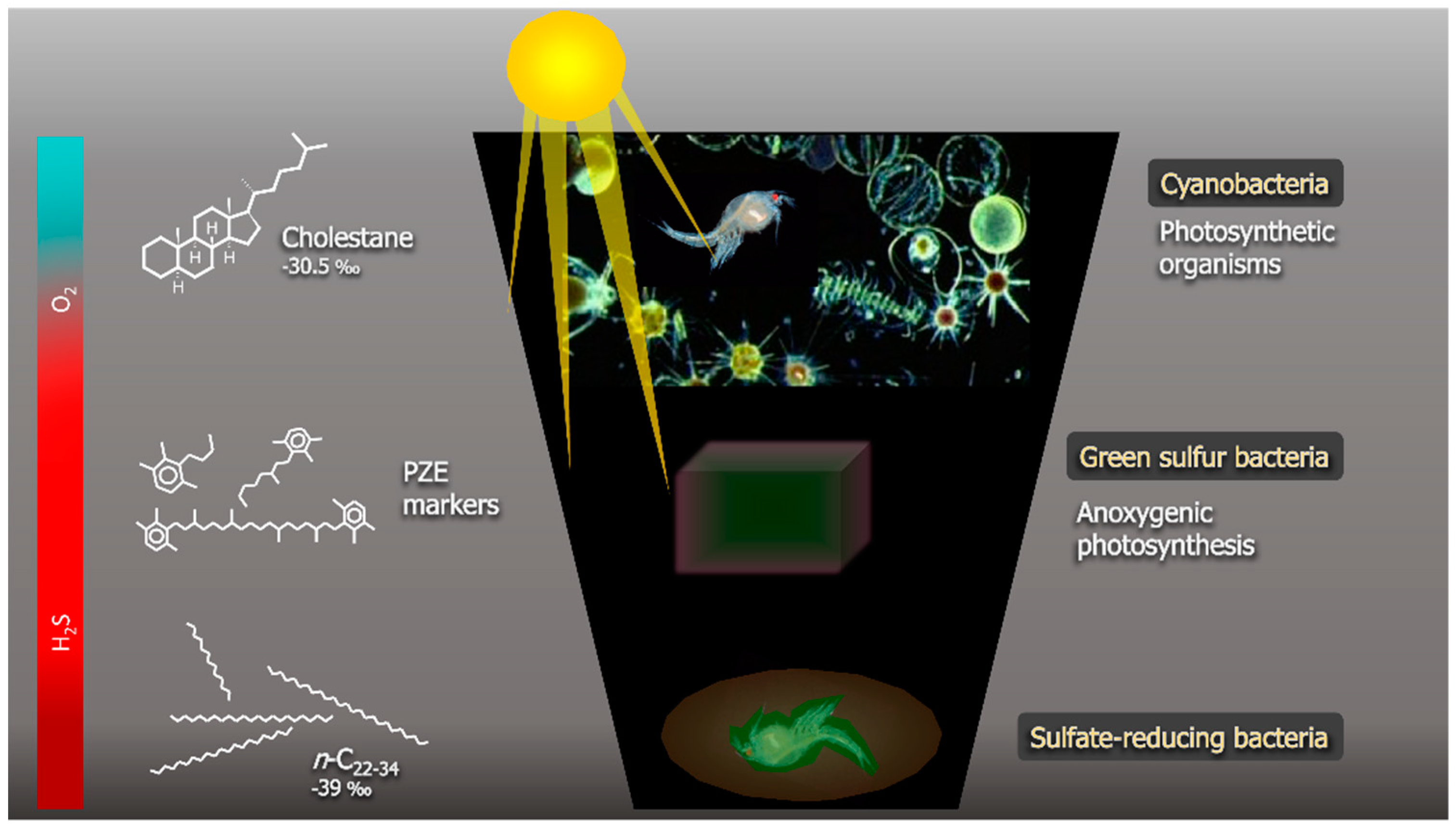
Conditions of photic zone euxinia (PZE), in which sulfidic (euxinic) waters extend in to the photic zone, have been shown to contribute to the excellent preservation of lipids in Phanerozoic marine environments [10,35,60,99]. PZE results from the enhanced activity of sulfate-reducing bacteria producing abundant reduced sulfur species (e.g., hydrogen sulfide) [102] which extends into the photic zone. Under these conditions, photosynthetic bacteria such as Chlorobi thrive [103]. These bacteria utilise unique diaromatic carotenoids during photosynthesis, and so, the diagenetic breakdown products of these carotenoids are highly specific markers for PZE in the sedimentary record [60,104,105].
In a study of a carbonate concretion from the Gogo Formation in the Canning Basin of Western Australia containing exceptionally preserved soft tissue of an invertebrate, Melendez et al. detected abundant biomarkers of Chlorobi, indicating that PZE conditions played a major role in the preservation of the soft tissues [10] (Figure 3). A distinctive distribution of steroid diagenetic products was also detected in this concretion with a predominance of C27 sterane [9]. Carbon isotope analysis (δ13C) and the abundance of cholestane indicated that the fossilised invertebrate was a crustacean, demonstrating the potential of exceptionally preserved fossils to contribute to chemotaxonomy [10]. A further study of small nonfossiliferous Gogo concretions and the surrounding host sediment revealed few differences which were suggested to be lithology-related [99].
The significance of biomarkers from encapsulated organic matter was further investigated in a study of concretions from the Lower Toarcian Posidonia Shale Formation (approx. 183 Ma) by Plet et al. [35]. In that study, the author studied the similarities in biomarker distribution between concretions and host sediment and highlighted the potential of carbonate concretions as palaeoenvironmental recorders. The indicators of photic zone anoxia and euxinia were reported. Moreover, the 13C-depletion of the carbonate and n-alkanes in the concretion compared to the host sediment indicated that sulfate-reducing bacteria facilitated concretion growth. Elevated hydrogen indices (HI) in the inner concretion compared to the outer concretion revealed that the organic matter preservation decreased towards the concretion margins [35].
Concretions containing exceptionally preserved fossilised soft tissue are ideal targets in the search for other labile biomolecules such as sterols, which are membrane lipids typically found in eukaryotes. Sterols and their diagenetic breakdown products are a highly useful group of biomarkers, as many organisms produce specific sterol distributions through which they can be identified in the sedimentary record [106,107]. The Gogo concretion studied by Melendez et al. [10] contained the earliest known occurrence of intact sterols directly associated with the encapsulated fossil crustacean, along with a full range of diagenetic products [9]. Despite its Devonian age (approx. 380 Ma), this sample yielded a complex and complete steroid continuum revealing the oldest preserved intact biological sterols alongside highly mature counterparts such as triaromatic steroids. [9] The biogeochemistry of fossils preserved within a carbonate concretion was further investigated on a Toarcian ichthyosaur vertebra [8]. This bone revealed the presence of collagen fibres and blood cell-like structures, i.e., red blood cells, white blood cells and platelets. Along with the preservation of these fragile features, the bone also preserved an abundance of cholesterol, with a stable isotopic composition confirming the ichthyosaur’s rank in the palaeo-food-chain [8].
Overall, recent studies have underlined that the investigation of biomarkers associated with fossils preserved in carbonate concretions contributes to unravelling the secrets of past ecosystems and environments. The isolated environment created by microbial ecosystem during concretion growth has the potential to provide crucial information to the understanding of the evolution of life throughout geological time.
6. Biomarkers in Concretions vs. Host Sediments
As carbonate concretions are more resistant to erosion and weathering, they are sometimes preserved where their host sediment has weathered away. Thus, understanding the palaeoenvironmental conditions associated with their formation is challenging. To determine if the biomarkers preserved in carbonate concretions can be used for palaeoenvironmental reconstruction, studies investigated carbonate concretions for which the host sediment was preserved. When compared, the host sediment and concretion show a strong resemblance in their biomarker composition. As most studies aimed to improve the understanding of the microbial processes and communities involved in the formation of the carbonate concretions, they have targeted the changes in biomarker distribution within concretions. Only a few studies have compared the biomarker composition of the concretion with that of the host rock.
Studies of Jurassic and Devonian concretions revealed that the distribution of saturated hydrocarbons is highly similar in the concretion and the host sediment [35,99]. Only minor variations were identified. In the Jurassic concretions, n-alkanes were consistently more depleted in 13C by approx. 1‰ than in the surrounding sediment; this depletion was attributed to the enhanced microbial activity leading to the concretion formation [35]. Lengger et al. [99] reported a small difference in the lithology-related biomarkers as well as a slightly greater C27 sterane and a lower 3-methylhopane contribution to the concretions compared to the host sediment. In both of these case studies, carbonate concretions were highly similar to their host sediments, which indicate that biomarker distribution within the carbonate concretions can be used for palaeoenvironmental reconstruction when the host sediment has been lost. In addition, in some instances where the biomarker signal of the concretion is superimposed on that of the host sediment, details of the microbial communities involved in the concretion formation can be revealed [108].
However, caution must be taken when averaging the concretion signals as some biomarker variations within a single concretion can occur and hold crucial information on the type of decaying organic matter around which the enhanced microbial activity took place [8,9,10]. As such, the averaging of concretion signals to reconstruct palaeoenvironments could lose the information unique to a particular concretion.
7. Conclusions
In “exceptional” fossil deposits (Konservat-Lagerstätten), well-preserved fossils formed via rapid preservation can provide significant insights into the history of organisms and their evolution. Konservat-Lagerstätten may also contain information on the palaeoenvironment and palaeoecological associations. Exceptional preservation can only occur under very specific circumstances, such as rapid burial, negligible bioturbation, anoxic or euxinic conditions and rapid entombment by microbial communities. The combination of some or all of these conditions can encourage rapid preservation and protect the organic matter from degradation. In particular, the microbially-initiated formation of carbonate concretions promotes the exceptional preservation of soft tissue. Concretions have been found throughout the geological record, and recent studies have investigated the exceptionally preserved biomolecules, biomarkers and other labile organic structures found within. These lipids and fossils preserved in concretions for hundreds of millions of years have provided insight into palaeobiology and the palaeoenvironment at the time of concretion formation. Biomarkers preserved in concretions are able to be traced back to their biological precursors and, thus, are critical in understanding the palaeoenvironment of deposition and preservation, as well as aspects of palaeobiology. A range of imaging techniques, as well as biomolecular analyses, are often used to investigate concretions. Concretions continue to reveal fossils with exceptional preservation of soft tissues, biomolecules and biomarkers, and these exciting findings may be the key to unravelling many evolutionary questions.
Author Contributions
Conceptualisation, K.G.; Writing-Original Draft Preparation, Review & Editing, K.G., A.I.H, C.P. and M.T.
Funding
K.G. acknowledges the Australian Research Council for a Discovery Outstanding Research Award (ARC DP130100577) and an ARC Infrastructure grant (ARC LE11010119).
Acknowledgments
Special thanks to Victor O. Leshyk (http://www.victorleshyk.com/) for the preparation of Figure 1. The authors thank the two anonymous reviewers for their thoughtful comments.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the writing of the manuscript or in the decision to publish.
References
- Briggs, D.E.G.; Crowther, P.R. Palaeobiology: A Synthesis; Blackwell Scientific Publications: Oxford, UK, 1990. [Google Scholar]
- Bottjer, D.J.; Etter, W.; Hagadorn, J.W.; Tang, C.M. Fossil-Lagerstätten: Jewels of the Fossil Record. In Exceptional Fossil Preservation: A Unique View on the Evolution of Marine Life; Bottjer, D.J., Etter, W., Hagadorn, J.W., Tang, C.M., Eds.; Columbia University Press: New York, NY, USA, 2002; pp. 1–10. [Google Scholar]
- Butterfield, N.J. Exceptional Fossil Preservation and the Cambrian Explosion. Integr. Comp. Biol. 2003, 43, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Greenwalt, D.E.; Goreva, Y.S.; Siljeström, S.M.; Rose, T.; Harbach, R.E. Hemoglobin-derived porphyrins preserved in a Middle Eocene blood-engorged mosquito. Proc. Natl Acad. Sci. USA 2013, 110, 18496–18500. [Google Scholar] [CrossRef] [PubMed]
- Bertazzo, S.; Maidment, S.C.R.; Kallepitis, C.; Fearn, S.; Stevens, M.M.; Xie, H.-n. Fibres and cellular structures preserved in 75-million–year-old dinosaur specimens. Nat. Commun. 2015, 6, 7352. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-C.; Chiang, C.-C.; Huang, P.-Y.; Chung, C.-Y.; Huang, T.D.; Wang, C.-C.; Chen, C.-I.; Chang, R.-S.; Liao, C.-H.; Reisz, R.R. Evidence of preserved collagen in an Early Jurassic sauropodomorph dinosaur revealed by synchrotron FTIR microspectroscopy. Nat. Commun. 2017, 8, 14220. [Google Scholar] [CrossRef] [PubMed]
- Wiemann, J.; Fabbri, M.; Yang, T.-R.; Stein, K.; Sander, P.M.; Norell, M.A.; Briggs, D.E.G. Fossilization transforms vertebrate hard tissue proteins into N-heterocyclic polymers. Nat. Commun. 2018, 9, 4741. [Google Scholar] [CrossRef] [PubMed]
- Plet, C.; Grice, K.; Pagès, A.; Verrall, M.; Coolen, M.J.L.; Ruebsam, W.; Rickard, W.D.A.; Schwark, L. Palaeobiology of red and white blood cell-like structures, collagen and cholesterol in an ichthyosaur bone. Sci. Rep. 2017, 7, 13776. [Google Scholar] [CrossRef] [PubMed]
- Melendez, I.; Grice, K.; Schwark, L. Exceptional preservation of Palaeozoic steroids in a diagenetic continuum. Sci. Rep. 2013, 3, 2768. [Google Scholar] [CrossRef] [PubMed]
- Melendez, I.; Grice, K.; Trinajstic, K.; Ladjavardi, M.; Greenwood, P.; Thompson, K. Biomarkers reveal the role of photic zone euxinia in exceptional fossil preservation: An organic geochemical perspective. Geology 2013, 41, 123–126. [Google Scholar] [CrossRef]
- Cohen, P.A.; Macdonald, F.A. The Proterozoic Record of Eukaryotes. Paleobiology 2015, 41, 610–632. [Google Scholar] [CrossRef]
- Muscente, A.D.; Schiffbauer, J.D.; Broce, J.; Laflamme, M.; O’Donnell, K.; Boag, T.H.; Meyer, M.; Hawkins, A.D.; Huntley, J.W.; McNamara, M.; et al. Exceptionally preserved fossil assemblages through geologic time and space. Gondwana Res. 2017, 48, 164–188. [Google Scholar] [CrossRef]
- Briggs, D.E.G. The role of decay and mineralization in the preservation of soft-bodied fossils. Annu. Rev. Earth Planet. Sci. 2003, 31, 275–301. [Google Scholar] [CrossRef]
- Declet, A.; Reyes, E.; Suárez, O.M. Calcium carbonate precipitation: A review of the carbonate crystallization process and applications in bioinspired composites. Rev. Adv. Mater. Sci. 2016, 44, 87–107. [Google Scholar]
- Rice, C.M.; Trewin, N.H.; Anderson, L.I. Geological setting of the Early Devonian Rhynie cherts, Aberdeenshire, Scotland: An early terrestrial hot spring system. J. Geol. Soc. Lond. 2002, 159, 203–214. [Google Scholar] [CrossRef]
- Sigleo, A.C. Organic geochemistry of silicified wood, Petrified Forest National Park, Arizona. Geochim. Cosmochim. Acta 1978, 42, 1397–1405. [Google Scholar] [CrossRef]
- Pawlowska, M.M.; Butterfield, N.J.; Brocks, J.J. Lipid taphonomy in the Proterozoic and the effect of microbial mats on biomarker preservation. Geology 2013, 41, 103–106. [Google Scholar] [CrossRef]
- Seilacher, A. Begriff und Bedeutung der Fossil-Lagerstätten. Neues Jahrbuch für Geologie und Paläontologie 1970, 1, 34–39. [Google Scholar]
- Parry, L.A.; Smithwick, F.; Nordén, K.K.; Saitta, E.T.; Lozano-Fernandez, J.; Tanner, A.R.; Caron, J.-B.; Edgecombe, G.D.; Briggs, D.E.G.; Vinther, J. Soft-Bodied Fossils Are Not Simply Rotten Carcasses—Toward a Holistic Understanding of Exceptional Fossil Preservation. BioEssays 2018, 40, 1700167. [Google Scholar] [CrossRef] [PubMed]
- Allison, P.A. Konservat-Lagerstaetten; cause and classification. Paleobiology 1988, 14, 331–344. [Google Scholar] [CrossRef]
- Gould, S.J. Wonderful Life: The Burgess Shale and the Nature of History; W. W. Norton & Company: New York, NY, USA, 1990. [Google Scholar]
- Selden, P.A.; Nudds, J.R. Evolution of Fossil Ecosystems; Manson Publishing: London, UK, 2012. [Google Scholar]
- Bobrovskiy, I.; Hope, J.M.; Ivantsov, A.; Nettersheim, B.J.; Hallmann, C.; Brocks, J.J. Ancient steroids establish the Ediacaran fossil Dickinsonia as one of the earliest animals. Science 2018, 361, 1246–1249. [Google Scholar] [CrossRef] [PubMed]
- Bobrovskiy, I.; Hope, J.M.; Krasnova, A.; Ivantsov, A.; Brocks, J.J. Molecular fossils from organically preserved Ediacara biota reveal cyanobacterial origin for Beltanelliformis. Nat. Ecol. Evol. 2018, 2, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Lerosey-Aubril, R.; Gaines, R.R.; Hegna, T.A.; Ortega-Hernández, J.; Van Roy, P.; Kier, C.; Bonino, E. The Weeks Formation Konservat-Lagerstätte and the evolutionary transition of Cambrian marine life. J. Geol. Soc. 2018, 175, 705–715. [Google Scholar] [CrossRef]
- Siveter, D.J. Ostracods in the Palaeozoic? Senckenbergiana lethaea 2008, 88, 1–9. [Google Scholar] [CrossRef]
- Siveter, D.J.; Aitchison, J.C.; Siveter, D.J.; Sutton, M.D. The Radiolaria of the Herefordshire Konservat-Lagerstätte (Silurian), England. J. Micropalaeontol. 2007, 26, 87–95. [Google Scholar] [CrossRef]
- Long, J.A.; Trinajstic, K. The Late Devonian Gogo Formation Lägerstatte of Western Australia: Exceptional Early Vertebrate Preservation and Diversity. Annu. Rev. Earth Planet. Sci. 2010, 38, 255–279. [Google Scholar] [CrossRef]
- Conway Morris, S.; Savoy, L.E.; Harris, A.G. An enigmatic organism from the "Exshaw" Formation (Devonian-Carboniferous), Alberta, Canada. Lethaia 1991, 24, 139–152. [Google Scholar] [CrossRef]
- Smith, M.G.; Bustin, R.M. Late Devonian and Early Mississippian Bakken and Exshaw Black Shale Source Rocks, Western Canada Sedimentary Basin: A Sequence Stratigraphic Interpretation. AAPG Bull. 2000, 84, 940–960. [Google Scholar] [CrossRef]
- Baird, G.C.; Sroka, S.D.; Shabica, C.W.; Kuecher, G.J. Taphonomy of Middle Pennsylvanian Mazon Creek area fossil localities, Northeast Illinois; significance of exceptional fossil preservation in syngenetic concretions. PALAIOS 1986, 1, 271–285. [Google Scholar] [CrossRef]
- Clements, T.; Purnell, M.; Gabbott, S. The Mazon Creek Lagerstätte: A diverse late Paleozoic ecosystem entombed within siderite concretions. J. Geol. Soc. 2019, 176, 1–11. [Google Scholar] [CrossRef]
- McCoy, V.E.; Young, R.T.; Briggs, D.E.G. Factors controlling exceptional preservation in concretions. Palaios 2015, 30, 272–280. [Google Scholar] [CrossRef]
- Fernandes, A.S. A Geobiological Investigation of the Mazon Creek Concretions of Northeastern Illinois, Mechanisms of Formation and Diagenesis. Master’s Thesis, The University of Western Ontario, London, ON, Canada, 2012. [Google Scholar]
- Plet, C.; Grice, K.; Pagès, A.; Ruebsam, W.; Coolen, M.J.L.; Schwark, L. Microbially-mediated fossil-bearing carbonate concretions and their significance for palaeoenvironmental reconstructions: A multi-proxy organic and inorganic geochemical appraisal. Chem. Geol. 2016, 426, 95–108. [Google Scholar] [CrossRef]
- Röhl, H.-J.; Schmid-Röhl, A.; Oschmann, W.; Frimmel, A.; Schwark, L. The Posidonia Shale (Lower Toarcian) of SW-Germany: An oxygen-depleted ecosystem controlled by sea level and palaeoclimate. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2001, 165, 27–52. [Google Scholar] [CrossRef]
- Brito, P.M.; Gallo, V. A new species of Lepidotes (Neopterygii: Semionotiformes: Semionotidae) from the Santana Formation, Lower Cretaceous of northeastern Brazil. J. Vertebr. Paleontol. 2003, 23, 47–53. [Google Scholar] [CrossRef]
- Fara, E.; Saraiva, A.Á.F.; de Almeida Campos, D.; Moreira, J.K.R.; de Carvalho Siebra, D.; Kellner, A.W.A. Controlled excavations in the Romualdo Member of the Santana Formation (Early Cretaceous, Araripe Basin, northeastern Brazil): Stratigraphic, palaeoenvironmental and palaeoecological implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 218, 145–160. [Google Scholar] [CrossRef]
- Heimhofer, U.; Hesselbo, S.P.; Pancost, R.D.; Martill, D.M.; Hochuli, P.A.; Guzzo, J.V.P. Evidence for photic-zone euxinia in the Early Albian Santana Formation (Araripe Basin, NE Brazil). Terra Nova 2008, 20, 347–354. [Google Scholar] [CrossRef]
- Rust, J. Fossil record of mass moth migration. Nature 2000, 405, 530–531. [Google Scholar] [CrossRef] [PubMed]
- Bertelli, S.; Lindow, B.E.K.; Dyke, G.J.; Chiappe, L.M. A well-preserved "charadriiform-like" fossil bird from the Early Eocene Fur Formation of Denmark. Palaeontology 2010, 53, 507–531. [Google Scholar] [CrossRef]
- Chen, P.-j.; Dong, Z.-m.; Zhen, S.-N. An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. Nature 1998, 391, 147–152. [Google Scholar] [CrossRef]
- Yang, W.; Li, S.; Jiang, B. New evidence for Cretaceous age of the feathered dinosaurs of Liaoning: Zircon U-Pb SHRIMP dating of the Yixian Formation in Sihetun, northeast China. Cretac. Res. 2007, 28, 177–182. [Google Scholar] [CrossRef]
- Mutterlose, J.; Böckel, B. The Barremian–Aptian interval in NW Germany: A review. Cretac. Res. 1998, 19, 539–568. [Google Scholar] [CrossRef]
- Mutterlose, J.; Pauly, S.; Steuber, T. Temperature controlled deposition of early Cretaceous (Barremian–early Aptian) black shales in an epicontinental sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 273, 330–345. [Google Scholar] [CrossRef]
- Ferber, C.T.; Wells, N.A. Paleolimnology and taphonomy of some fish deposits in “Fossil” and “Uinta” Lakes of the Eocene Green River Formation, Utah and Wyoming. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1995, 117, 185–210. [Google Scholar] [CrossRef]
- Conrad, J.L.; Head, J.J.; Carrano, M.T. Unusual Soft-Tissue Preservation of a Crocodile Lizard (Squamata, Shinisauria) From the Green River Formation (Eocene) and Shinisaur Relationships. Anat. Rec. 2014, 297, 545–559. [Google Scholar] [CrossRef] [PubMed]
- Tissot, B.P.; Welte, D.H. Petroleum Formation and Occurence; Springer: Berlin, Germany, 1984. [Google Scholar]
- Nip, M.; Tegelaar, E.W.; Brinkhuis, H.; De Leeuw, J.W.; Schenck, P.A.; Holloway, P.J. Analysis of modern and fossil plant cuticles by Curie point Py-GC and Curie point Py-GC-MS: Recognition of a new, highly aliphatic and resistant biopolymer. Org. Geochem. 1986, 10, 769–778. [Google Scholar] [CrossRef]
- Goth, K.; de Leeuw, J.W.; Püttmann, W.; Tegelaar, E.W. Origin of Messel Oil Shale kerogen. Nature 1988, 336, 759–761. [Google Scholar] [CrossRef]
- Gelin, F.; de Leeuw, J.W.; Sinninghe Damsté, J.S.; Derenne, S.; Largeau, C.; Metzger, P. The similarity of chemical structures of soluble aliphatic polyaldehyde and insoluble algaenan in the green microalga Botryococcus braunii race A as revealed by analytical pyrolysis. Org. Geochem. 1994, 21, 423–435. [Google Scholar] [CrossRef]
- Grice, K.; Audino, M.; Boreham, C.J.; Alexander, R.; Kagi, R.I. Distributions and stable carbon isotopic compositions of biomarkers in torbanites from different palaeogeographical locations. Org. Geochem. 2001, 32, 1195–1210. [Google Scholar] [CrossRef]
- Grice, K.; Schouten, S.; Blokker, P.; Derenne, S.; Largeau, C.; Nissenbaum, A.; Damsté, J.S.S. Structural and isotopic analysis of kerogens in sediments rich in free sulfurised Botryococcus braunii biomarkers. Org. Geochem. 2003, 34, 471–482. [Google Scholar] [CrossRef]
- McCoy, V.E. Concretions as Agents of Soft-Tissue Preservation: A Review. Paleontol. Soc. Pap. 2014, 20, 147–162. [Google Scholar] [CrossRef]
- Marshall, J.D.; Pirrie, D. Carbonate concretions—explained. Geol. Today 2013, 29, 53–62. [Google Scholar] [CrossRef]
- Kiriakoulakis, K.; Marshall, J.D.; Wolff, G.A. Biomarkers in a Lower Jurassic concretion from Dorset (UK). J. Geol. Soc. 2000, 157, 207–220. [Google Scholar] [CrossRef]
- Gaines, R.R.; Vorhies, J.S. Growth mechanisms and geochemistry of carbonate concretions from the Cambrian Wheeler Formation (Utah, USA). Sedimentology 2016, 63, 662–698. [Google Scholar] [CrossRef]
- Zhu, T.; Dittrich, M. Carbonate Precipitation through Microbial Activities in Natural Environment, and Their Potential in Biotechnology: A Review. Front. Bioeng. Biotechnol. 2016, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Kremer, B.; Kazmierczak, J.; Stal, l.J. Calcium carbonate precipitation in cyanobacterial mats from sandy tidal flats of the North Sea. Geobiology 2008, 6, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Schwark, L. Exceptional preservation of microbial lipids in Paleozoic to Mesoproterozoic sediments. Geology 2013, 41, 287–288. [Google Scholar] [CrossRef]
- Coleman, M.L. Microbial processes: Controls on the shape and composition of carbonate concretions. Mar. Geol. 1993, 113, 127–140. [Google Scholar] [CrossRef]
- Raiswell, R.; Fisher, Q.J. Mudrock-hosted carbonate concretions: A review of growth mechanisms and their influence on chemical and isotopic composition. J. Geol. Soc. 2000, 157, 239–251. [Google Scholar] [CrossRef]
- Pye, K.; Dickson, J.A.D.; Schiavon, N.; Coleman, M.L.; Cox, M. Formation of siderite-Mg-calcite-iron sulphide concretions in intertidal marsh and sandflat sediments, north Norfolk, England. Sedimentology 1990, 37, 325–343. [Google Scholar] [CrossRef]
- Curtis, C.D.; Coleman, M.L.; Love, L.G. Pore water evolution during sediment burial from isotopic and mineral chemistry of calcite, dolomite and siderite concretions. Geochim. Cosmochim. Acta 1986, 50, 2321–2334. [Google Scholar] [CrossRef]
- Berner, R.A. Sulphate Reduction, Organic Matter Decomposition and Pyrite Formation. Phil. Trans. R. Soc. Lond. Ser. A Math. Phys. Sci. 1985, 315, 25–38. [Google Scholar] [CrossRef]
- Woodland, B.G.; Stenstrom, R.C. The occurrence and origin of siderite concretions in the Francis Creek Shale (Pennsylvanian) of northeastern Illinois. In Mazon Creek Fossils; Nitecki, M.H., Ed.; Academic Press: Cambridge, MA, USA, 1979; pp. 69–103. [Google Scholar]
- Curtis, C.D.; Spears, D.A. The formation of sedimentary iron minerals. Econ. Geol. 1968, 63, 257–270. [Google Scholar] [CrossRef]
- Ho, C.; Coleman, J.M. Consolidation and Cementation of Recent Sediments in the Atchafalaya Basin. GSA Bull. 1969, 80, 183–192. [Google Scholar] [CrossRef]
- Pye, K. SEM analysis of siderite cements in intertidal marsh sediments, Norfolk, England. Mar. Geol. 1984, 56, 1–12. [Google Scholar] [CrossRef]
- Brocks, J.J.; Pearson, A. Building the Biomarker Tree of Life. Rev. Mineral. Geochem. 2005, 59, 233–258. [Google Scholar] [CrossRef]
- Volkman, J.K. Lipid Markers for Marine Organic Matter. In Marine Organic Matter: Biomarkers, Isotopes and DNA; Volkman, J.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Pearson, A. Lipidomics for Geochemistry. In Treatise on Geochemistry (Second Edition) Volume 12: Organic Geochemistry; Falkowski, P.G., Freeman, K.H., Eds.; Elsevier: Oxford, UK, 2014; pp. 291–336. [Google Scholar]
- Maixner, F.; Overath, T.; Linke, D.; Janko, M.; Guerriero, G.; van den Berg, B.H.J.; Stade, B.; Leidinger, P.; Backes, C.; Jaremek, M.; et al. Paleoproteomic study of the Iceman’s brain tissue. Cell. Mol. Life Sci. 2013, 70, 3709–3722. [Google Scholar] [CrossRef] [PubMed]
- More, K.D.; Orsi, W.D.; Galy, V.; Giosan, L.; He, L.; Grice, K.; Coolen, M.J.L. A 43 kyr record of protist communities and their response to oxygen minimum zone variability in the Northeastern Arabian Sea. Earth Planet. Sci. Lett. 2018, 496, 248–256. [Google Scholar] [CrossRef]
- Orsi, W.D.; Coolen, M.J.L.; Wuchter, C.; He, L.; More, K.D.; Irigoien, X.; Chust, G.; Johnson, C.; Hemingway, J.D.; Lee, M.; et al. Climate oscillations reflected within the microbiome of Arabian Sea sediments. Sci. Rep. 2017, 7, 6040. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, J.A.; Rahman, I.A.; Lautenschlager, S.; Rayfield, E.J.; Donoghue, P.C.J. A virtual world of paleontology. Trends Ecol. Evol. 2014, 29, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Mietchen, D.; Aberhan, M.; Manz, B.; Hampe, O.; Mohr, B.; Neumann, C.; Volke, F. Three-dimensional Magnetic Resonance Imaging of fossils across taxa. Biogeosciences 2008, 5, 25–41. [Google Scholar] [CrossRef]
- Holgado, B.; Dalla Vecchia, F.M.; Fortuny, J.; Bernardini, F.; Tuniz, C. A Reappraisal of the Purported Gastric Pellet with Pterosaurian Bones from the Upper Triassic of Italy. PLOS ONE 2015, 10, e0141275. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.D.; Brett, C.E. Concretions as sources of exceptional preservation, and decay as a source of concretions: Examples from the Middle Devonian of New York. PALAIOS 2013, 28, 305–316. [Google Scholar] [CrossRef]
- Pradel, A.; Langer, M.; Maisey, J.G.; Geffard-Kuriyama, D.; Cloetens, P.; Janvier, P.; Tafforeau, P. Skull and brain of a 300-million-year-old chimaeroid fish revealed by synchrotron holotomography. Proc. Natl. Acad. Sci. USA 2009, 106, 5224–5228. [Google Scholar] [CrossRef] [PubMed]
- Garwood, R.J.; Dunlop, J.A.; Selden, P.A.; Spencer, A.R.T.; Atwood, R.C.; Vo, N.T.; Drakopoulos, M. Almost a spider: A 305-million-year-old fossil arachnid and spider origins. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160125. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.; Ahlberg, P.E.; Trinajstic, K.M.; Mirone, A.; Tafforeau, P. Three-Dimensional Synchrotron Virtual Paleohistology: A New Insight into the World of Fossil Bone Microstructures. Microsc. Microanal. 2012, 18, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Sutton, M.D. Tomographic techniques for the study of exceptionally preserved fossils. Proc. R. Soc. B Biol. Sci. 2008, 275, 1587–1593. [Google Scholar] [CrossRef] [PubMed]
- Sutton, M.D.; Rahman, I.A.; Garwood, R.J. Techniques for Virtual Palaeontology; John Wiley & Sons, Ltd.: Chichester, UK, 2014. [Google Scholar]
- Siveter, D.J.; Briggs, D.E.G.; Siveter, D.J.; Sutton, M.D.; Legg, D. A new crustacean from the Herefordshire (Silurian) Lagerstätte, UK, and its significance in malacostracan evolution. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170279. [Google Scholar] [CrossRef] [PubMed]
- Bertsch, P.M.; Hunter, D.B. Applications of Synchrotron-Based X-ray Microprobes. Chem. Rev. 2001, 101, 1809–1842. [Google Scholar] [CrossRef] [PubMed]
- Mozley, P.S. Complex compositional zonation in concretionary siderite; implications for geochemical studies. J. Sediment. Res. 1989, 59, 815–818. [Google Scholar] [CrossRef]
- De Craen, M.; Swennen, R.; Keppens, E. Petrography and geochemistry of septarian carbonate concretions from the Boom Clay Formation (Oligocene, Belgium). Geol. Mijnb. 1998, 77, 63–76. [Google Scholar] [CrossRef]
- Danise, S.; Twitchett, R.J.; Matts, K. Ecological succession of a Jurassic shallow-water ichthyosaur fall. Nat. Commun. 2014, 5, 4789. [Google Scholar] [CrossRef] [PubMed]
- Vickerman, J.C. ToF-SIMS—An Overview. In ToF-SIMS: Surface Analysis by Mass Spectrometry; Vickerman, J.C., Briggs, D., Eds.; IM Publications: Chichester, UK, 2001; pp. 1–36. [Google Scholar]
- Thiel, V.; Sjövall, P. Using Time-of-Flight Secondary Ion Mass Spectrometry to Study Biomarkers. Annu. Rev. Earth Planet. Sci. 2011, 39, 125–156. [Google Scholar] [CrossRef]
- Abbott, G.D.; Fletcher, I.W.; Tardio, S.; Hack, E. Exploring the geochemical distribution of organic carbon in early land plants: A novel approach. Phil. Trans. R. Soc. B Biol. Sci. 2017, 373, 20160499. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, M.H.; Zheng, W.; Organ, C.L.; Avci, R.; Suo, Z.; Freimark, L.M.; Lebleu, V.S.; Duncan, M.B.; Vander Heiden, M.G.; Neveu, J.M.; et al. Biomolecular Characterization and Protein Sequences of the Campanian Hadrosaur B. canadensis. Science 2009, 324, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Thiel, V.; Blumenberg, M.; Kiel, S.; Leefmann, T.; Liebenau, K.; Lindgren, J.; Sjövall, P.; Treude, T.; Zilla, T. Occurrence and fate of fatty acyl biomarkers in an ancient whale bone (Oligocene, El Cien Formation, Mexico). Org. Geochem. 2014, 68, 71–81. [Google Scholar] [CrossRef]
- Lindgren, J.; Uvdal, P.; Sjövall, P.; Nilsson, D.E.; Engdahl, A.; Schultz, B.P.; Thiel, V. Molecular preservation of the pigment melanin in fossil melanosomes. Nat. Commun. 2012, 3, 824. [Google Scholar] [CrossRef] [PubMed]
- Colleary, C.; Dolocan, A.; Gardner, J.; Singh, S.; Wuttke, M.; Rabenstein, R.; Habersetzer, J.; Schaal, S.; Feseha, M.; Clemens, M.; et al. Chemical, experimental, and morphological evidence for diagenetically altered melanin in exceptionally preserved fossils. Proc. Natl. Acad. Sci. USA 2015, 112, 12592–12597. [Google Scholar] [CrossRef] [PubMed]
- Moyer, A.E.; Zheng, W.; Johnson, E.A.; Lamanna, M.C.; Li, D.-Q.; Lacovara, K.J.; Schweitzer, M.H. Melanosomes or Microbes: Testing an Alternative Hypothesis for the Origin of Microbodies in Fossil Feathers. Sci. Rep. 2014, 4, 4233. [Google Scholar] [CrossRef] [PubMed]
- Grice, K.; Eiserbeck, C. The Analysis and Application of Biomarkers. In Treatise on Geochemistry (Second Edition) Volume 12: Organic Geochemistry; Falkowski, P.G., Freeman, K.H., Eds.; Elsevier: Oxford, UK, 2014; pp. 47–78. [Google Scholar]
- Lengger, S.K.; Melendez, I.M.; Summons, R.E.; Grice, K. Mudstones and embedded concretions show differences in lithology-related, but not source-related biomarker distributions. Org. Geochem. 2017, 113, 67–74. [Google Scholar] [CrossRef]
- Briggs, D.E.G.; Summons, R.E. Ancient biomolecules: Their origins, fossilization, and role in revealing the history of life. BioEssays 2014, 36, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, A.J.M.; Cox, G.M.; Brocks, J.J.; Grosjean, E.; Boreham, C.J.; Edwards, D.S. Microbial assemblage and palaeoenvironmental reconstruction of the 1.38 Ga Velkerri Formation, McArthur Basin, northern Australia. Geobiology 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- Canfield, D.E.; Farquhar, J. The global sulfur cycle. In Fundamentals of Geobiology; Knoll, A.H., Canfield, D.E., Konhauser, K.O., Eds.; Blackwell Publishing Ltd.: Chichester, UK, 2012; pp. 49–64. [Google Scholar]
- Grice, K.; Cao, C.; Love, G.D.; Böttcher, M.E.; Twitchett, R.J.; Grosjean, E.; Summons, R.E.; Turgeon, S.C.; Dunning, W.; Jin, Y. Photic zone euxinia during the Permian-Triassic superanoxic event. Science 2005, 307, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Grice, K.; Schaeffer, P.; Schwark, L.; Maxwell, J.R. Molecular indicators of palaeoenvironmental conditions in an immature Permian shale (Kupferschiefer, Lower Rhine Basin, north-west Germany) from free and S-bound lipids. Org. Geochem. 1996, 25, 131–147. [Google Scholar] [CrossRef]
- Summons, R.E.; Powell, T.G. Chlorobiaceae in Palaeozoic seas revealed by biological markers, isotopes and geology. Nature 1986, 319, 763–765. [Google Scholar] [CrossRef]
- Volkman, J.K. A review of sterol markers for marine and terrigenous organic matter. Org. Geochem. 1986, 9, 83–99. [Google Scholar] [CrossRef]
- Volkman, J.K. Sterols in microorganisms. Appl. Microbiol. Biotechnol. 2003, 60, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Thiel, V.; Hoppert, M. Fatty acids and other biomarkers in two Early Jurassic concretions and their immediate host rocks (Lias δ, Buttenheim clay pit, Bavaria, Germany). Org. Geochem. 2018, 120, 42–55. [Google Scholar] [CrossRef]
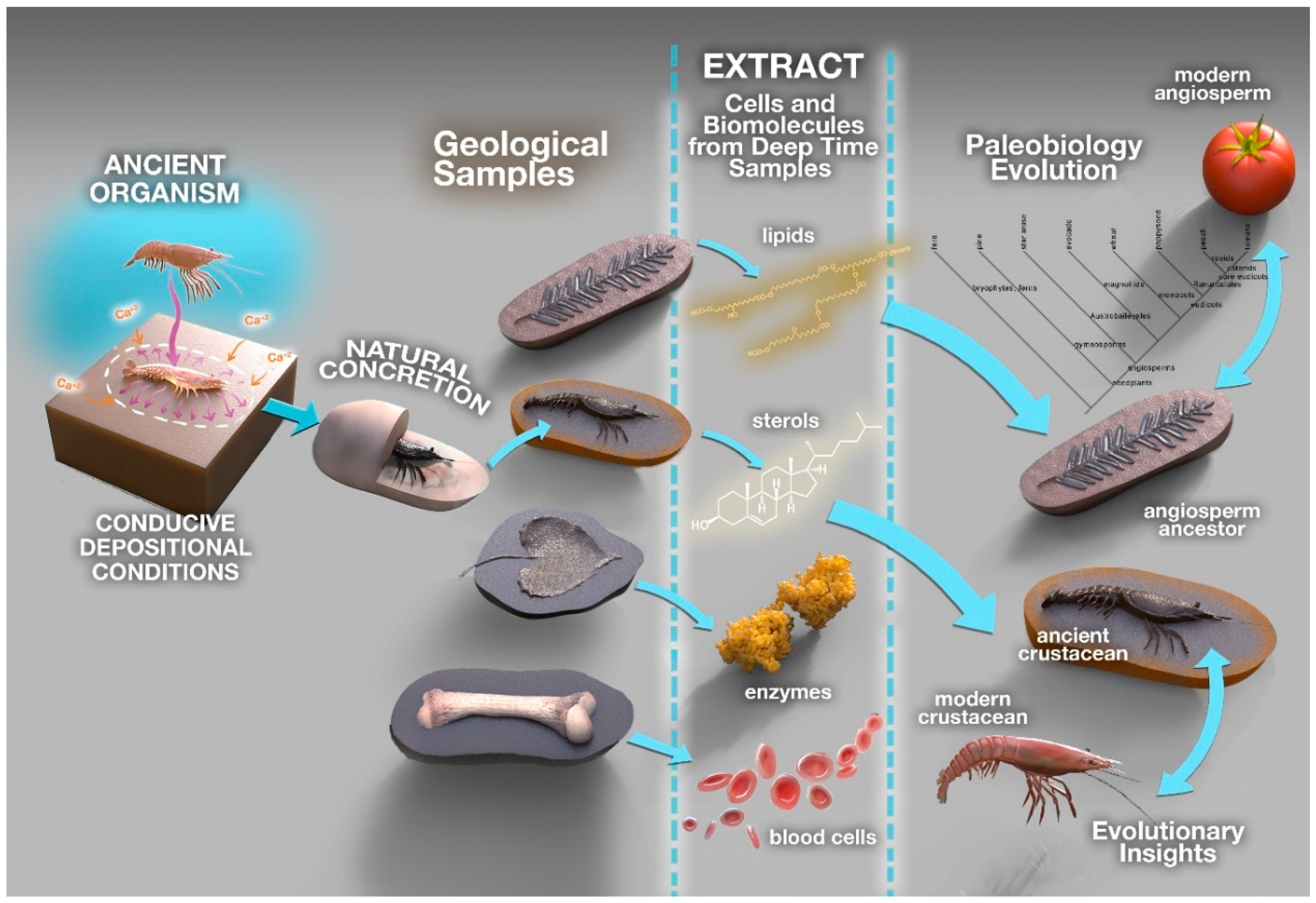
Figure 1.
Studies of exceptionally preserved fossils provide information on the links between modern and ancient organisms, e.g., References [8,10,34].

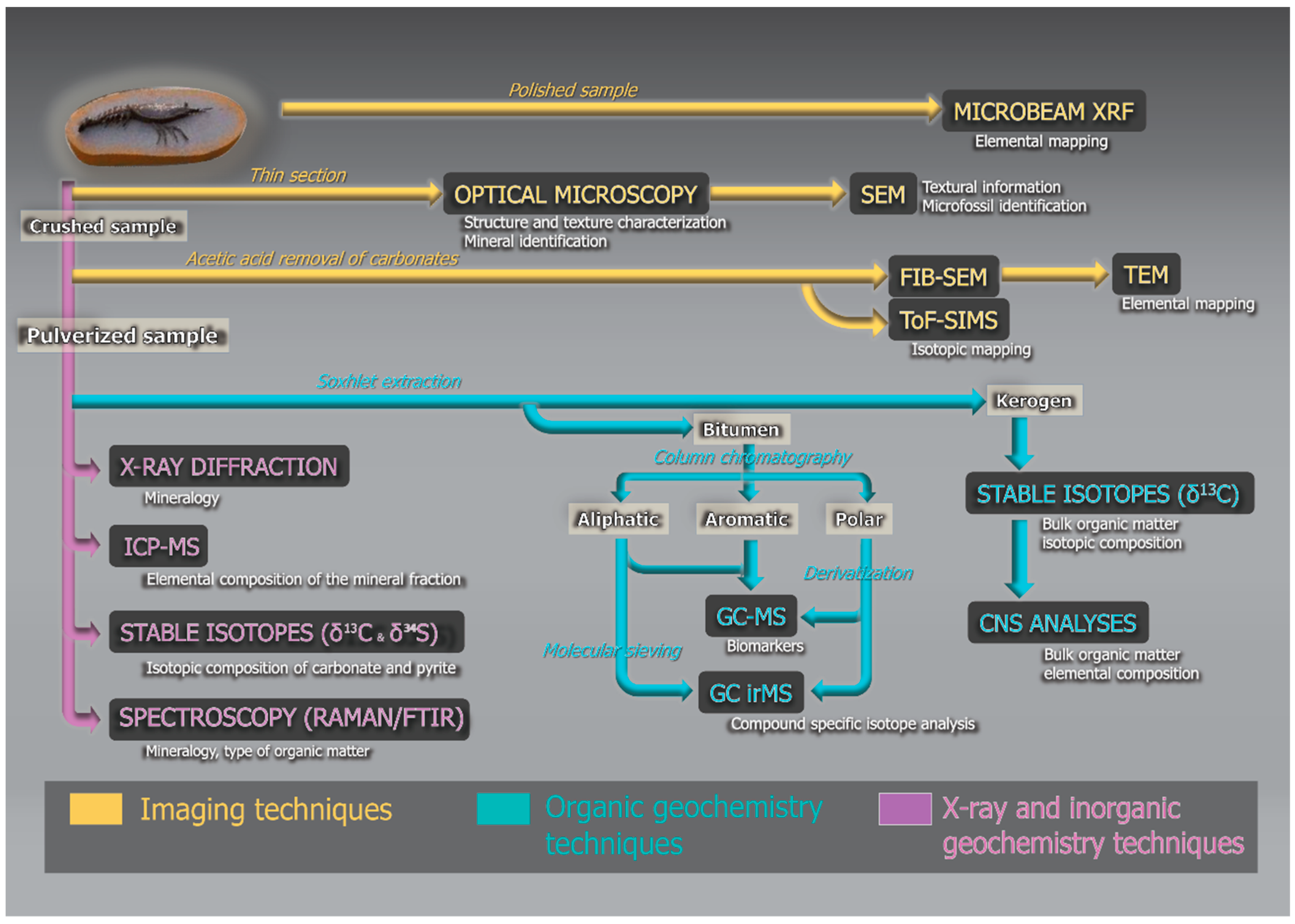
Figure 2.
A typical analytical flowchart for the analysis of exceptionally preserved fossils, including nondestructive imaging techniques, and organic and inorganic geochemistry.
Figure 2.
A typical analytical flowchart for the analysis of exceptionally preserved fossils, including nondestructive imaging techniques, and organic and inorganic geochemistry.

Figure 3.
The preservation mechanism of a fossilised crustacean inside a carbonate concretion, as described by Melendez et al. [10].
Figure 3.
The preservation mechanism of a fossilised crustacean inside a carbonate concretion, as described by Melendez et al. [10].


{kind=link}
{kind=link}
{kind=link}
Table 1.
The locations of notable Konservat-Lagerstätten and Konzentrat-Lagerstätten samples.
| Site | Age | Description | References |
|---|---|---|---|
| White Sea (Russia) | Ediacaran | Exceptional preservation of Ediacaran biota including Beltanelliformis and Dickinsonia | [23,24] |
| Burgess Shale (British Columbia) | Cambrian | Well-preserved soft tissues from marine arthropod groups (e.g., trilobites and crustaceans); role of cyanobacterial mats in preservation and extinction | [21] |
| Weeks Fm (Utah, USA) | Cambrian | Diverse fauna: animal community living in open marine distal shelf environment | [25] |
| Herefordshire (UK) | Silurian | Radiolarians: worms, molluscs, starfish, brachiopods, arthropods and other fossils of unknown affinity | [26,27] |
| Gogo Fm (Western Australia) | Devonian | Three-dimensional preservation of fish (including placoderms), crustaceans and other marine organisms | [9,10,28] |
| Exshaw Fm (Western Canada) | Devonian | Preserved soft-body fossils of uncertain assignment, potentially hydrozoan | [29,30] |
| Mazon Creek (Illinois) | Carboniferous | Terrestrial and aquatic flora and fauna including extinct seed ferns; early evolution, fire and stress markers | [31,32,33,34] |
| Posidonia Shale (Southwest Germany) | Early Jurassic | Exceptionally well-preserved marine fossil skeletons including ichthyosaurs, ammonites and crinoids; adaptation to environment, e.g., low O2 levels (blood cell morphology of ichthyosaurs) | [8,35,36] |
| Santana Fm (Brazil) | Cretaceous | Assemblages of fossilised flora, arthropods, insects, fish, turtles, snakes and dinosaurs | [37,38,39] |
| Fur Fm (Denmark) | Cretaceous | Remains of birds, reptiles, fish, insects, crustaceans, molluscs and diatoms | [40,41] |
| Yixian Fm (China) | Cretaceous | Well-preserved feathered dinosaurs and birds, along with a selection of non-theropod dinosaurs | [42,43] |
| Blätterton Fm (Germany) | Cretaceous | Carbonate, siderite and phosphorate concretions, all of which have been shown to yield fossils | [44,45] |
| Green River Fm (Wyoming) | Eocene | Fish, birds, insects and leaves | [46,47] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Grice, K.; Holman, A.I.; Plet, C.; Tripp, M. Fossilised Biomolecules and Biomarkers in Carbonate Concretions from Konservat-Lagerstätten. Minerals 2019, 9, 158. https://doi.org/10.3390/min9030158
AMA Style
Grice K, Holman AI, Plet C, Tripp M. Fossilised Biomolecules and Biomarkers in Carbonate Concretions from Konservat-Lagerstätten. Minerals. 2019; 9(3):158. https://doi.org/10.3390/min9030158
Chicago/Turabian StyleGrice, Kliti, Alex I. Holman, Chloe Plet, and Madison Tripp. 2019. "Fossilised Biomolecules and Biomarkers in Carbonate Concretions from Konservat-Lagerstätten" Minerals 9, no. 3: 158. https://doi.org/10.3390/min9030158
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.