Pushing the Limits of Seagrass Remote Sensing in the Turbid Waters of Elkhorn Slough, California

 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
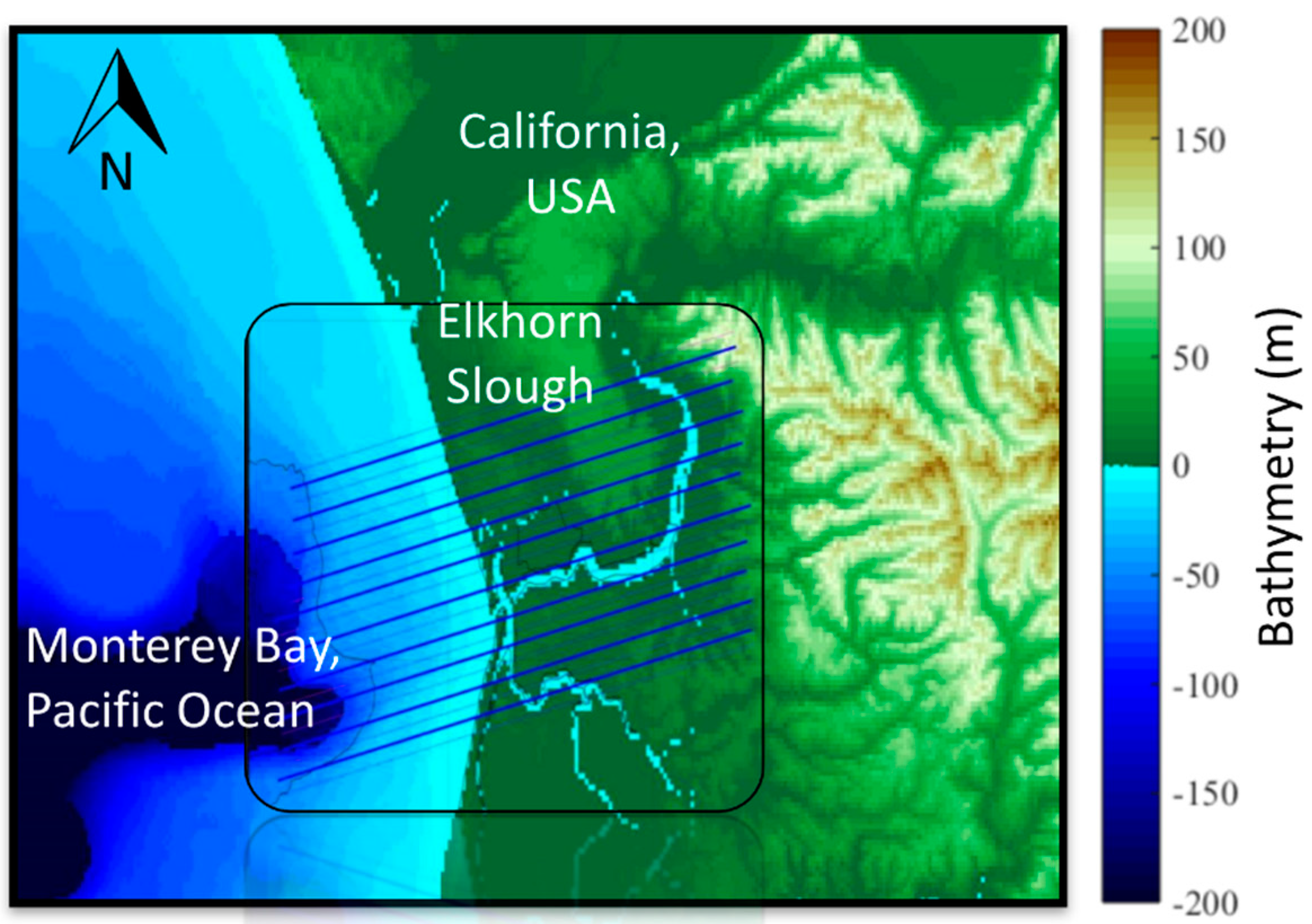
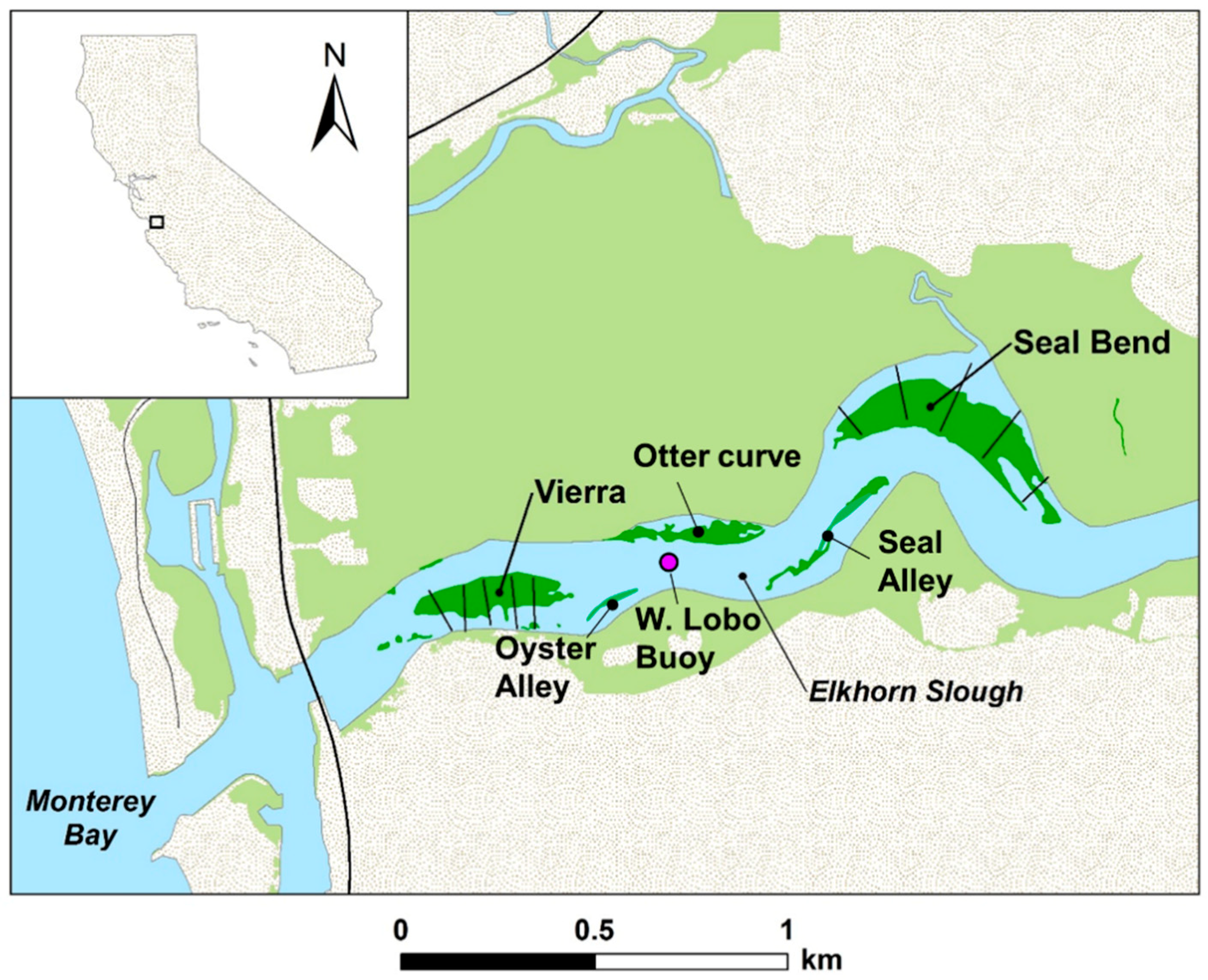
2.1. Study Location
2.2. Field Data
2.3. Google Earth and Sentinel 2 Imagery
2.4. Hyperspectral Airborne Imagery
2.5. Benthic Retrieval Algorithms
2.5.1. Red Edge Retrieval
2.5.2. Semi-Analytical Inversion Method
3. Results and Discussion
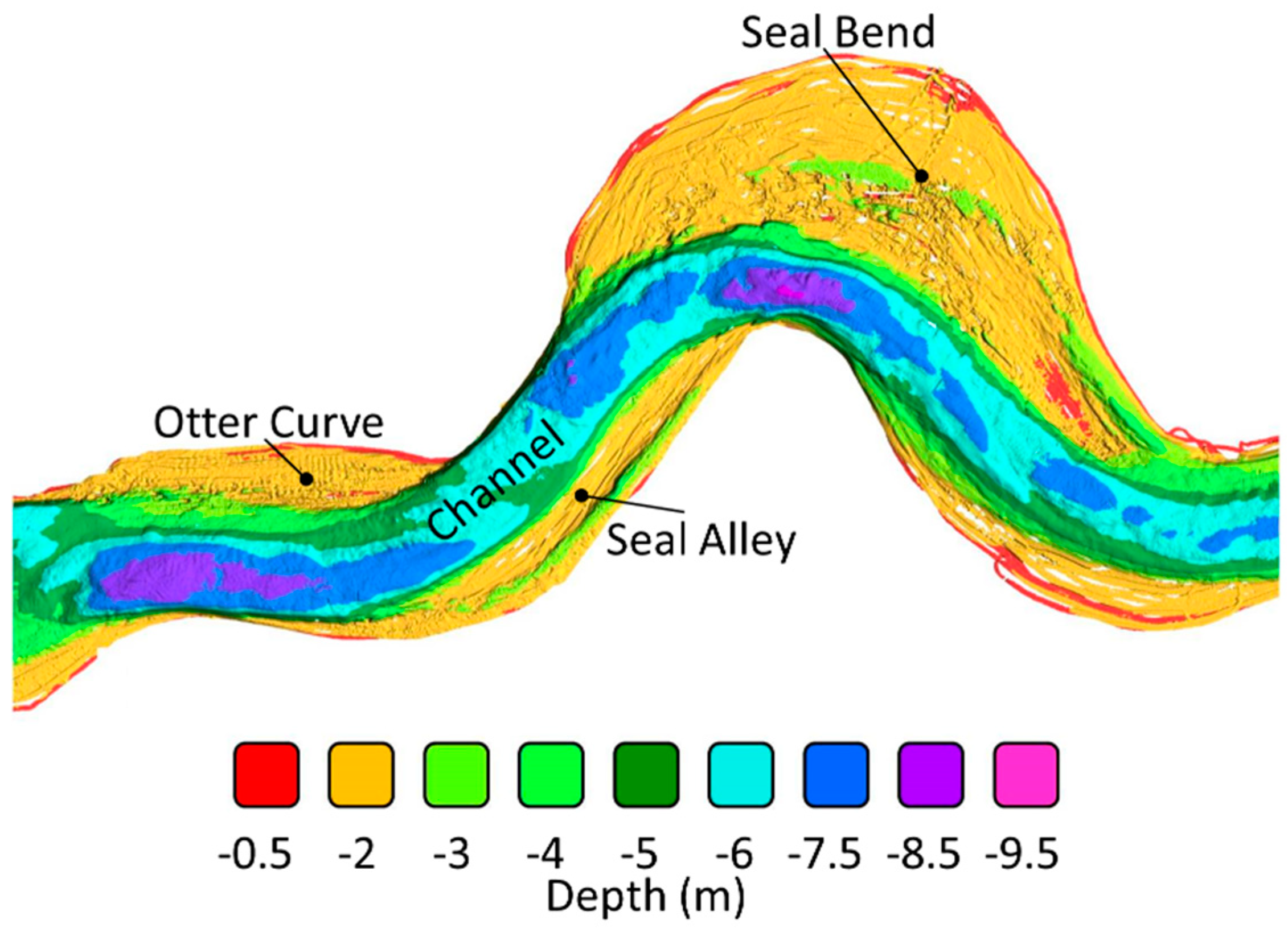
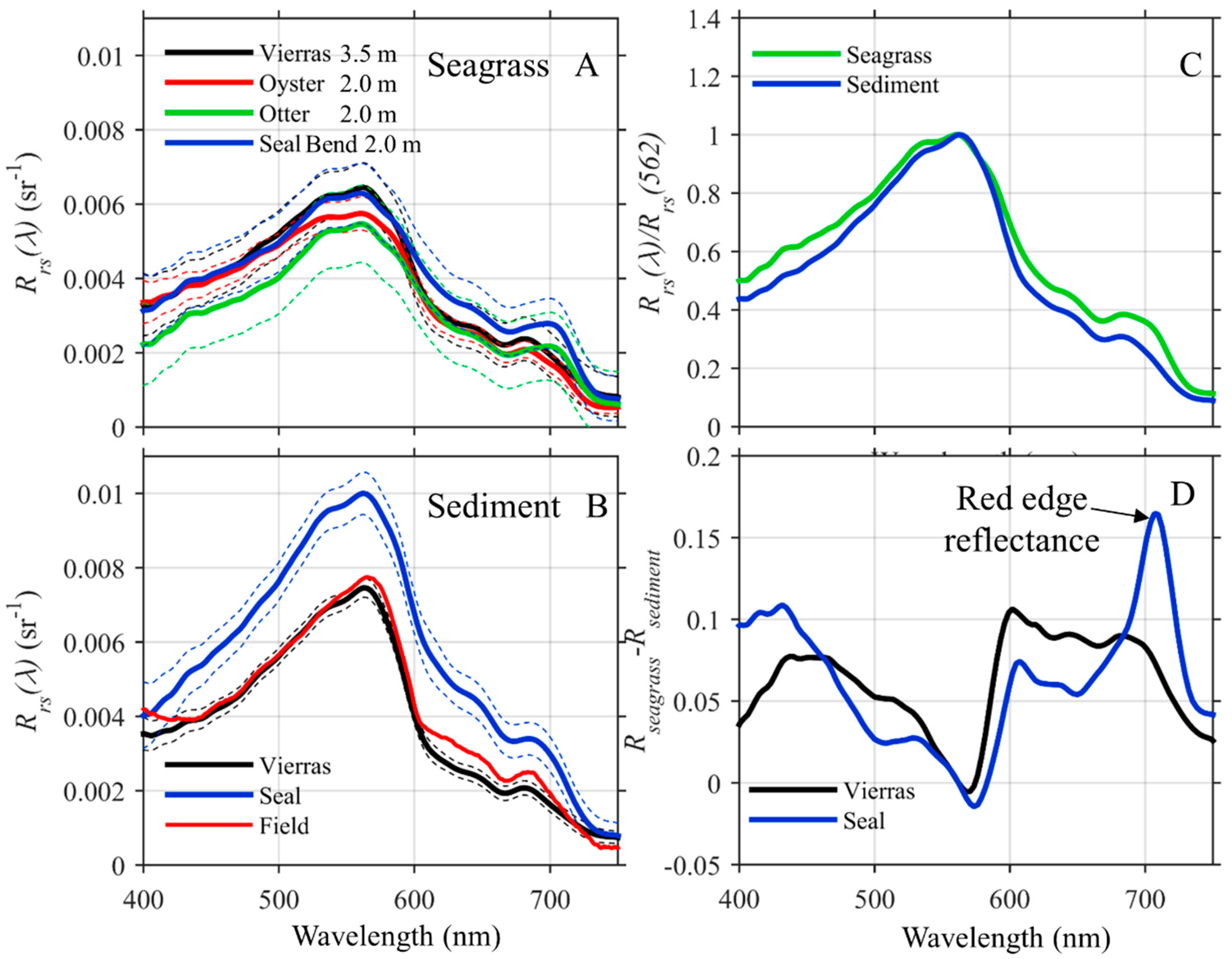
3.1. Field Characterization of the Benthos
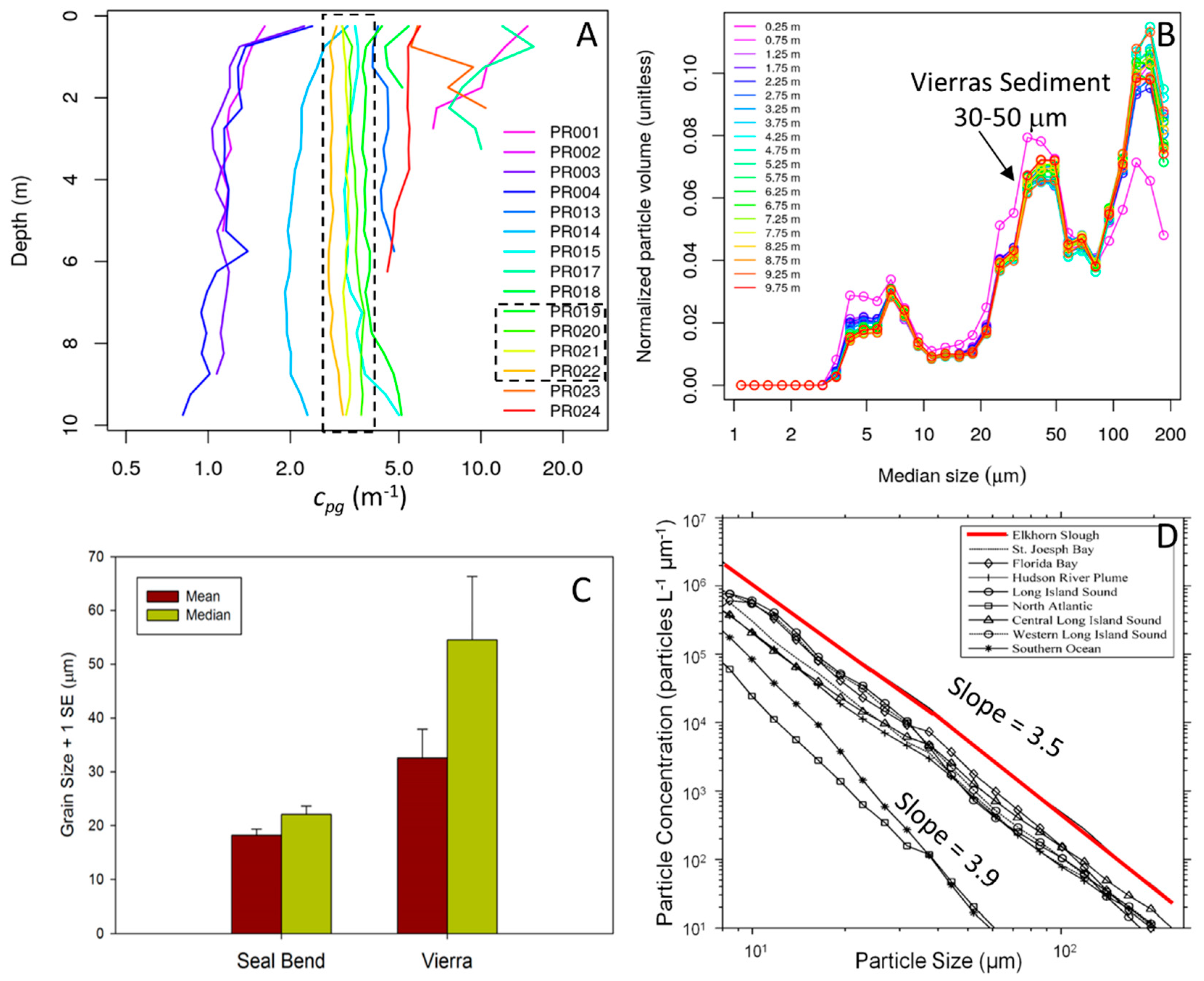
3.2. Optical Properties of Seafloor and Water Column
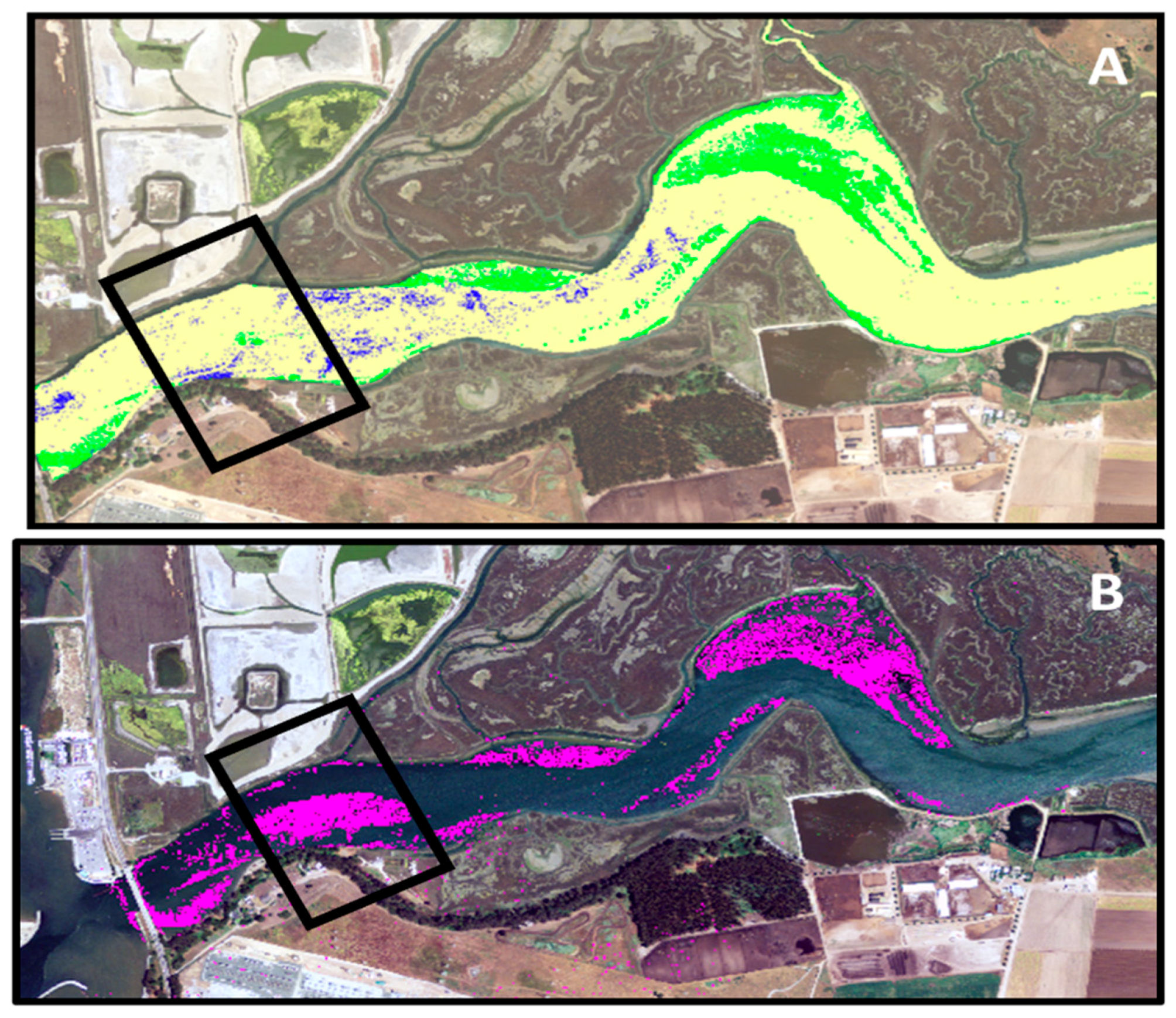
3.3. Google Earth Imagery
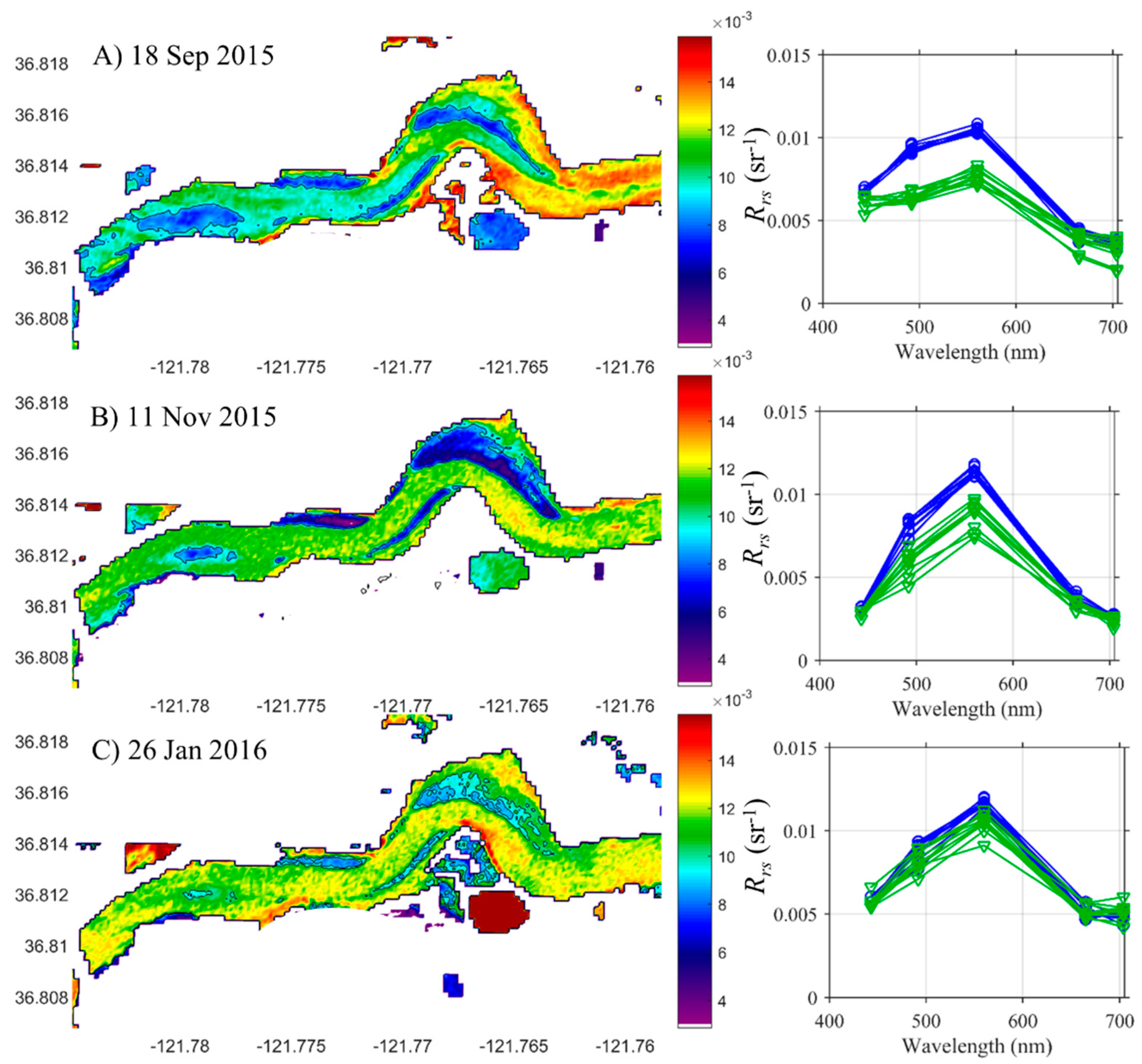
3.4. Satellite and Aircraft Spectrometry
3.4.1. Red Edge and Threshold Algorithms
3.4.2. Semi-Analytical Inversion Modeling
4. Conclusions and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dierssen, H.M.; Zimmerman, R.C.; Leathers, R.A.; Downes, T.V.; Davis, C.O. Ocean color remote sensing of seagrass and bathymetry in the Bahamas Banks by high resolution airborne imagery. Limnol. Oceanogr. 2003, 48, 444–455. [Google Scholar] [CrossRef]
- Dekker, A.G.; Brando, V.E.; Anstee, J.M. Retrospective seagrass change detection in a shallow coastal tidal Australian lake. Remote Sens. Environ. 2005, 97, 415–433. [Google Scholar] [CrossRef]
- Dierssen, H.M. Benthic ecology from space: Optics and net primary production in seagrass and benthic algae across the Great Bahama Bank. Mar. Ecol. Prog. Ser. 2010, 411, 1–15. [Google Scholar] [CrossRef]
- Hedley, J.; Russell, B.; Randolph, K.; Dierssen, H. A physics-based method for the remote sensing of seagrasses. Remote Sens. Environ. 2016, 174, 134–147. [Google Scholar] [CrossRef]
- Ackleson, S.G.; Klemas, V. Remote sensing of submerged vegetation in lower Chesapeake Bay: A comparison of Landsat MSS to TM Imagery. Remote Sens. Environ. 1987, 22, 235–248. [Google Scholar] [CrossRef]
- Phinn, S.; Roelfsema, C.; Dekker, A.; Brando, V.; Anstee, J. Mapping seagrass species, cover and biomass in shallow waters: An assessment of satellite multi-spectral and airborne hyper-spectral imaging systems in Moreton Bay (Australia). Remote Sens. Environ. 2008, 112, 3413–3425. [Google Scholar] [CrossRef]
- Wabnitz, C.C.; Andréfouët, S.; Torres-Pulliza, D.; Müller-Karger, F.E.; Kramer, P.A. Regional-scale seagrass habitat mapping in the Wider Caribbean region using Landsat sensors: Applications to conservation and ecology. Remote Sens. Environ. 2008, 112, 3455–3467. [Google Scholar] [CrossRef]
- Lyons, M.B.; Phinn, S.R.; Roelfsema, C.M. Long term land cover and seagrass mapping using Landsat and object-based image analysis from 1972 to 2010 in the coastal environment of South East Queensland, Australia. ISPRS J. Photogramm. Remote Sens. 2012, 71, 34–46. [Google Scholar] [CrossRef]
- Pu, R.; Bell, S.; Meyer, C.; Baggett, L.; Zhao, Y. Mapping and assessing seagrass along the western coast of Florida using Landsat TM and EO-1 ALI/Hyperion imagery. Estuar. Coast. Shelf Sci. 2012, 115, 234–245. [Google Scholar] [CrossRef]
- Dekker, A.G.; Phinn, S.R.; Anstee, J.; Bissett, P.; Brando, V.E.; Casey, B.; Fearns, P.; Hedley, J.; Klonowski, W.; Lee, Z.P.; et al. Intercomparison of shallow water bathymetry, hydro-optics, and benthos mapping techniques in Australian and Caribbean coastal environments. Limnol. Oceanogr. Methods 2011, 9, 396–425. [Google Scholar] [CrossRef] [Green Version]
- Hedley, J.; Russell, B.J.; Randolph, K.; Pérez-Castro, M.Á.; Vásquez-Elizondo, R.M.; Enríquez, S.; Dierssen, H.M. Remote sensing of seagrass leaf area index and species: The capability of a model inversion method assessed by sensitivity analysis and hyperspectral data of Florida Bay. Front. Mar. Sci. 2017, 4, 362. [Google Scholar] [CrossRef]
- Petit, T.; Bajjouk, T.; Mouquet, P.; Rochette, S.; Vozel, B.; Delacourt, C. Hyperspectral remote sensing of coral reefs by semi-analytical model inversion—Comparison of different inversion setups. Remote Sens. Environ. 2017, 190, 348–365. [Google Scholar] [CrossRef]
- Hochberg, E.J.; Atkinson, M.J. Capabilities of remote sensors to classify coral, algae, and sand as pure and mixed spectra. Remote Sens. Environ. 2003, 85, 174–189. [Google Scholar] [CrossRef]
- Kutser, T.; Vahtmäe, E.; Metsamaa, L. Spectral library of macroalgae and benthic substrates in Estonian coastal waters. Proc. Est. Acad. Sci. Biol. Ecol. 2006, 55, 329–340. [Google Scholar]
- Vahtmae, E.; Kutser, T.; Martin, G.; Kotta, J. Feasibility of hyperspectral remote sensing for mapping benthic macroalgal cover in turbid coastal waters—A Baltic Sea case study. Remote Sens. Environ. 2006, 101, 342–351. [Google Scholar] [CrossRef]
- Broenkow, W.W.; Breaker, L. A 30-year History of Tide and Current Measurements in Elkhorn Slough, California; Moss Landing Marine Laboratories: Moss Landing, CA, USA, 2005. [Google Scholar]
- Hughes, B.B.; Haskins, J.C.; Wasson, K.; Watson, E. Identifying factors that influence expression of eutrophication in a central California estuary. Mar. Ecol. Prog. Ser. 2011, 439, 31–43. [Google Scholar] [CrossRef]
- Hughes, B.B.; Eby, R.; Van Dyke, E.; Tinker, M.T.; Marks, C.I.; Johnson, K.S.; Wasson, K. Recovery of a top predator mediates negative eutrophic effects on seagrass. Proc. Natl. Acad. Sci. USA 2013, 110, 15313–15318. [Google Scholar] [CrossRef] [Green Version]
- Orth, R.J. Effect of nutrient enrichment on growth of the eelgrass Zostera marina in the Chesapeake Bay, Virginia, USA. Mar. Biol. 1977, 44, 187–194. [Google Scholar] [CrossRef]
- Tomasko, D.A.; Corbett, C.A.; Greening, H.S.; Raulerson, G.E. Spatial and temporal variation in seagrass coverage in Southwest Florida: Assessing the relative effects of anthropogenic nutrient load reductions and rainfall in four contiguous estuaries. Mar. Pollut. Bull. 2005, 50, 797–805. [Google Scholar] [CrossRef]
- Orth, R.J.; Carruthers, T.J.B.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L., Jr.; Hughes, A.R.; Kendrick, G.A.; Kenworthy, W.J.; Olyarnik, S.; et al. A global crisis for seagrass ecosystems. Bioscience 2006, 56, 987–996. [Google Scholar] [CrossRef]
- Byrd, K.B.; Kelly, N.M.; Van Dyke, E. Decadal changes in a Pacific estuary: A multi-source remote sensing approach for historical ecology. GIScience Remote Sens. 2004, 41, 347–370. [Google Scholar]
- Dierssen, H.M. Overview of hyperspectral remote sensing for mapping marine benthic habitats from airborne and underwater sensors. In Imaging Spectrometry XVIII; SPIE: San Diego, CA, USA, 2013; Volume 8870, pp. 1–7. [Google Scholar]
- Buonassissi, C.J.; Dierssen, H.M. A regional comparison of particle size distributions and the power law approximation in oceanic and estuarine surface waters. J. Geophys. Res. 2010, 115. [Google Scholar] [CrossRef]
- Vahtmäe, E.; Kutser, T. Classifying the Baltic Sea shallow water habitats using image-based and spectral library methods. Remote Sens. 2013, 5, 2451–2474. [Google Scholar] [CrossRef]
- Bostrom, K.J. Testing the Limits of Hyperspectral Airborne Remote Sensing by Mapping Eelgrass in Elkhorn Slough. Master’s Thesis, University of Connecticut, Mansfield, CT, USA, 2011. [Google Scholar]
- Mouroulis, P.; Green, R.O.; Chrien, T.G. Design of pushbroom imaging spectrometers for optimum recovery of spectroscopic and spatial information. Appl. Opt. 2000, 39, 2210–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouroulis, P.; Green, R.O.; Wilson, D.W. Optical design of a coastal ocean imaging spectrometer. Opt. Express 2008, 16, 9087–9096. [Google Scholar] [CrossRef] [PubMed]
- Mouroulis, P.; Van Gorp, B.; Green, R.O.; Dierssen, H.; Wilson, D.W.; Eastwood, M.; Boardman, J.; Gao, B.-C.; Cohen, D.; Franklin, B. The Portable Remote Imaging Spectrometer (PRISM) coastal ocean sensor: Design, characteristics and first flight results. Appl. Opt. 2013, 53, 1363–1380. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.R.; Gao, B.-C.; Green, R.O.; Roberts, D.A.; Dennison, P.E.; Lundeen, S.R. Atmospheric correction for global mapping spectroscopy: ATREM advances for the HyspIRI preparatory campaign. Remote Sens. Environ. 2015, 167, 64–77. [Google Scholar] [CrossRef]
- Thompson, D.R.; Seidel, F.C.; Gao, B.C.; Gierach, M.M.; Green, R.O.; Kudela, R.M.; Mouroulis, P. Optimizing irradiance estimates for coastal and inland water imaging spectroscopy. Geophys. Res. Lett. 2015, 42, 4116–4123. [Google Scholar] [CrossRef]
- Thompson, D.R.; Boardman, J.W.; Eastwood, M.L.; Green, R.O.; Haag, J.M.; Mouroulis, P.; Van Gorp, B. Imaging spectrometer stray spectral response: In-flight characterization, correction, and validation. Remote Sens. Environ. 2018, 204, 850–860. [Google Scholar] [CrossRef]
- Hill, V.J.; Zimmerman, R.C.; Bissett, W.P.; Dierssen, H.; Kohler, D.D. Evaluating Light Availability, Seagrass Biomass, and Productivity Using Hyperspectral Airborne Remote Sensing in Saint Joseph’s Bay, Florida. Estuaries Coasts 2014, 37, 1467–1489. [Google Scholar] [CrossRef]
- Zimmerman, R. Radiative transfer in seagrass canopies. Limnol. Oceanogr. 2003, 48, 568–585. [Google Scholar] [CrossRef]
- Hedley, J. A three-dimensional radiative transfer model for shallow water environments. Opt. Express 2008, 16, 21887–21902. [Google Scholar] [CrossRef] [PubMed]
- CEOS. Feasibility Study for an Aquatic Ecosystem Earth Observing System; Version 1.1; Dekker, A.G., Pinnel, N., Eds.; Committee on Earth Observation Satellites: Canberra, Australia, 2017. [Google Scholar]
- Muller-Karger, F.E.; Hestir, E.; Ade, C.; Turpie, K.; Roberts, D.A.; Siegel, D.; Miller, R.J.; Humm, D.; Izenberg, N.; Keller, M. Satellite sensor requirements for monitoring essential biodiversity variables of coastal ecosystems. Ecol. Appl. 2018, 28, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Chapin, T.P.; Caffrey, J.M.; Jannasch, H.W.; Coletti, L.J.; Haskins, J.C.; Johnson, K.S. Nitrate sources and sinks in Elkhorn Slough, California: Results from long-term continuous in situ nitrate analyzers. Estuaries 2004, 27, 882–894. [Google Scholar] [CrossRef]
- Dean, E.W. Tidal Scour in Elkhorn Slough, California: A Bathymetric Analysis; Monterey Bay; Faculty of Earth Systems Science & Policy, Center for Science, Technology and Information Resources, California State University: Monterey, CA, USA, 2003. [Google Scholar]
- Hammerstrom, K.; Grant, N. Assessment and Monitoring of Ecological Characteristics of Zostera Marina L Beds in Elkhorn Slough, California; Elkhorn Slough Technical Report Series 2012: 3; Elkhorn Slough Foundation: Moss Landing, CA, USA, 2012. [Google Scholar]
- Twardowski, M.S.; Donaghay, P.L. Separating in situ and terrigenous sources of absorption by dissolved materials in coastal waters. J. Geophys. Res. Ocean. (1978–2012) 2001, 106, 2545–2560. [Google Scholar] [CrossRef]
- Sullivan, J.M.; Twardowski, M.S.; Zaneveld, J.R.V.; Moore, C.M.; Barnard, A.H.; Donaghay, P.L.; Rhoades, B. Hyperspectral temperature and salt dependencies of absorption by water and heavy water in the 400–750 nm spectral range. Appl. Opt. 2006, 45, 5294–5309. [Google Scholar] [CrossRef] [PubMed]
- Zaneveld, J.R.V.; Moore, C.; Barnard, A.; Twardowski, M.S.; Chang, G.C. Correction and Analysis of Spectral Absorption Data Taken with the WET Labs AC-S; Office of Naval Research: Freemantle, Australia, 2004.
- Roesler, C.S.; Perry, M.J.; Carder, K.L. Modeling in situ phytoplankton absorption from total absorption spectra in productive inland marine waters. Limnol. Oceanogr. 1989, 34, 1510–1523. [Google Scholar] [CrossRef] [Green Version]
- Schofield, O.; Bergmann, T.; Oliver, M.J.; Irwin, A.; Kirkpatrick, G.; Bissett, W.P.; Moline, M.A.; Orrico, C. Inversion of spectral absorption in the optically complex coastal waters of the Mid-Atlantic Bight. J. Geophys. Res Ocean. 2004, 109. [Google Scholar] [CrossRef] [Green Version]
- Morel, A. Optical properties of pure water and pure seawater. In Optical Aspects of Oceanography; Academic: New York, NY, USA, 1974; pp. 1–24. [Google Scholar]
- Sullivan, J.M.; Twardowski, M.S.; Donaghay, P.L.; Freeman, S.A. Use of optical scattering to discriminate particle types in coastal waters. Appl. Opt. 2005, 44, 1667–1680. [Google Scholar] [CrossRef]
- Lee, Z.; Carder, K.L.; Mobley, C.D.; Steward, R.G.; Patch, J.S. Hyperspectral Remote Sensing for Shallow Waters. 2. Deriving Bottom Depths and Water Properties by Optimization. Appl. Opt. 1999, 38, 3831–3843. [Google Scholar] [CrossRef]
- Gould, R.W.; Arnone, R.A.; Sydor, M. Absorption, scattering, and remote sensing reflectance relationships in coastal waters: Testing a new inversion algorithm. J. Coast. Res. 2001, 17, 328–341. [Google Scholar]
- Moore, K.A. NERRS SWMP Bio-Monitoring Protocol: LONG-Term Monitoring of Estuarine Submersed and Emergent Vegetation Communities; National Estuarine Research Reserve System Technical Report; NOAA–NERRS: Silver Spring, MD, USA, 2009.
- Vanhellemont, Q. Adaptation of the dark spectrum fitting atmospheric correction for aquatic applications of the Landsat and Sentinel-2 archives. Remote Sens. Environ. 2019, 225, 175–192. [Google Scholar] [CrossRef]
- Gao, B.C.; Montes, M.J.; Ahmad, Z.; Davis, C.O. Atmospheric correction algorithm for hyperspectral remote sensing of ocean color from space. Appl. Opt. 2000, 39, 887–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermote, E.F.; Tanré, D.; Deuze, J.L.; Herman, M.; Morcette, J.-J. Second simulation of the satellite signal in the solar spectrum, 6S: An overview. IEEE Trans. Geosci. Remote Sens. 1997, 35, 675–686. [Google Scholar] [CrossRef]
- Hestir, E.L.; Khanna, S.; Andrew, M.E.; Santos, M.J.; Viers, J.H.; Greenberg, J.A.; Rajapakse, S.S.; Ustin, S.L. Identification of invasive vegetation using hyperspectral remote sensing in the California Delta ecosystem. Remote Sens. Environ. 2008, 112, 4034–4047. [Google Scholar] [CrossRef]
- Pope, R.; Fry, E. Absorption spectrum of pure water: 2. Integrating cavity measurements. Appl. Opt. 1997, 36, 8710–8723. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.R.; Hochberg, E.J.; Asner, G.P.; Green, R.O.; Knapp, D.E.; Gao, B.-C.; Garcia, R.; Gierach, M.; Lee, Z.; Maritorena, S. Airborne mapping of benthic reflectance spectra with Bayesian linear mixtures. Remote Sens. Environ. 2017, 200, 18–30. [Google Scholar] [CrossRef]
- Goodman, J.A.; Ustin, S.L. Classification of benthic composition in a coral reef environment using spectral unmixing. J. Appl. Remote Sens. 2007, 1, 011501. [Google Scholar]
- Klonowski, W.M.; Fearns, P.R.; Lynch, M.J. Retrieving key benthic cover types and bathymetry from hyperspectral imagery. J. Appl. Remote Sens. 2007, 1, 011505. [Google Scholar] [CrossRef]
- Garcia, R.; Lee, Z.; Hochberg, E. Hyperspectral Shallow-Water Remote Sensing with an Enhanced Benthic Classifier. Remote Sens. 2018, 10, 147. [Google Scholar] [CrossRef]
- McPherson, M.L.; Hill, V.J.; Zimmerman, R.C.; Dierssen, H.M. The optical properties of Greater Florida Bay: Implications for seagrass abundance. Estuaries Coasts 2011, 34, 1150–1160. [Google Scholar] [CrossRef]
- Thorhaug, A.; Richardson, A.D.; Berlyn, G.P. Spectral reflectance of the seagrasses: Thalassia testudinum, Halodule wrightii, Syringodium filiforme and five marine algae. Int. J. Remote Sens. 2007, 28, 1487–1501. [Google Scholar] [CrossRef]
- Gitelson, A. The peak near 700 nm on radiance spectra of algae and water: Relationships of its magnitude and position with chlorophyll concentration. Int. J. Remote Sens. 1992, 13, 3367–3373. [Google Scholar] [CrossRef]
- Dierssen, H.M.; Kudela, R.M.; Ryan, J.P.; Zimmerman, R.C. Red and black tides: Quantitative analysis of water-leaving radiance and perceived color for phytoplankton, colored dissolved organic matter, and suspended sediments. Limnol. Oceanogr. 2006, 51, 2646–2659. [Google Scholar] [CrossRef] [Green Version]
- Dierssen, H.M.; Zimmerman, R.C.; Burdige, D.J. Optics and remote sensing of Bahamian carbonate sediment whitings and potential relationship to wind-driven Langmuir circulation. Biogeosciences 2009, 6, 487–500. [Google Scholar] [CrossRef] [Green Version]
- Twardowski, M.S.; Boss, E.; Macdonald, J.B.; Pegau, W.S.; Barnard, A.H.; Zaneveld, J.R.V. A model for estimating bulk refractive index from the optical backscattering ratio and the implications for understanding particle composition in case I and case II waters. J. Geophys. Res. 2001, 105, 14129–14142. [Google Scholar] [CrossRef]
- Rottgers, R.; Doerffer, R.; McKee, D.; Schonfeld, W. Algorithm Theoretical Basis Document: The Water Optical Properties Processor (WOPP); Tech. Rep.; Helmholtz-Zentrum Geesthacht, University of Strathclyde: Geesthacht, Germany, 2011. [Google Scholar]
- Gambi, M.C.; Nowell, A.R.; Jumars, P.A. Flume observations on flow dynamics in Zostera marina (eelgrass) beds. Mar. Ecol. Prog. Ser. Oldendorf 1990, 61, 159–169. [Google Scholar] [CrossRef]
- Han, B.; Loisel, H.; Vantrepotte, V.; Mériaux, X.; Bryère, P.; Ouillon, S.; Dessailly, D.; Xing, Q.; Zhu, J. Development of a Semi-Analytical Algorithm for the Retrieval of Suspended Particulate Matter from Remote Sensing over Clear to Very Turbid Waters. Remote Sens. 2016, 8, 211. [Google Scholar] [CrossRef]
- Fogarty, M.C.; Fewings, M.R.; Paget, A.C.; Dierssen, H.M. The influence of a sandy substrate, seagrass, or highly turbid water on Albedo and surface heat flux. J. Geophys. Res. Ocean. 2018, 123, 53–73. [Google Scholar] [CrossRef]
- Campbell, J.B. Introduction to Remote Sensing, 2nd ed.; The Guilford Press: New York, NY, USA, 1996. [Google Scholar]
- Chirayath, V.; Earle, S.A. Drones that see through waves–preliminary results from airborne fluid lensing for centimetre-scale aquatic conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 237–250. [Google Scholar] [CrossRef]
- Hedley, J.; Mirhakak, M.; Wentworth, A.; Dierssen, H. Influence of Three-Dimensional Coral Structures on Hyperspectral Benthic Reflectance and Water-Leaving Reflectance. Appl. Sci. 2018, 8, 2688. [Google Scholar] [CrossRef]
- Joyce, K.E.; Phinn, S.R. Bi-directional reflectance of corals. Int. J. Remote Sens. 2002, 23, 389–394. [Google Scholar] [CrossRef]
- Hedley, J.; Enrıquez, S. Optical properties of canopies of the tropical seagrass Thalassia testudinum estimated by a three-dimensional radiative transfer model. Limnol. Oceanogr 2010, 55, 1537–1550. [Google Scholar] [CrossRef]
- Barnes, B.B.; Garcia, R.; Hu, C.; Lee, Z. Multi-band spectral matching inversion algorithm to derive water column properties in optically shallow waters: An optimization of parameterization. Remote Sens. Environ. 2018, 204, 424–438. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Google Earth Image Date | PRISM | |||
|---|---|---|---|---|---|
| 10/2011 | 05/2012 | 05/2013 | 08/2013 | 07/2012 | |
| Vierra | 0 1 | 0.5 | 0.5 | 1 | 1 |
| Oyster Alley | 0 | 0 | 0.5 | 1 | 1 |
| Otter Curve | 1 | 1 | 1 | 1 | 1 |
| Seal Alley | 0 | 0.5 | 1 | 0.5 | 1 |
| Seal Bend | 1 | 1 | 1 | 0.5 | 1 |
| Region | Indices | ||
|---|---|---|---|
| NDI(700, 670) | NDI(731.3, 673.6) | Rrs(675)/Rrs(550) | |
| Seagrass Threshold | >0 | >0.1 | >0.35 |
| Vierra | −0.0691 | 0.40 | 0.368 |
| Oyster Alley | −0.071 | 0.53 | 0.357 |
| Otter Curve | 0.057 | 0.42 | 0.370 |
| Seal Bend | 0.041 | 0.43 | 0.418 |
| Sediment Threshold | <0 | <0.1 | <0.35 |
| Vierra | −0.090 | 0.40 | 0.283 |
| Seal Bend | −0.059 | 0.54 | 0.342 |
| Parameter | PRISM Imagery | Field Data |
|---|---|---|
| P, aph(440) (m−1) | 0.15 ± 0.04 | 0.19 ± 0.041 |
| G, adg(440) (m−1) | 0.31 ± 0.07 | 0.37 ± 0.09 |
| X, bbp(550) (m−1) | 0.016 ± 0.005 | 0.059 ± 0.009 1 |
| H (Depth) (m) | 2.58 ± 1.21 | 1.96–4.46 2 |
| Chlorophyll a (mg·m−3) | 4.00 ± 1.92 | 4.75 ± 0.72 |
| TSM (g·m−3) | 1.02 ± 0.93 3 | 3.96 ± 0.33 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dierssen, H.M.; Bostrom, K.J.; Chlus, A.; Hammerstrom, K.; Thompson, D.R.; Lee, Z. Pushing the Limits of Seagrass Remote Sensing in the Turbid Waters of Elkhorn Slough, California. Remote Sens. 2019, 11, 1664. https://doi.org/10.3390/rs11141664
Dierssen HM, Bostrom KJ, Chlus A, Hammerstrom K, Thompson DR, Lee Z. Pushing the Limits of Seagrass Remote Sensing in the Turbid Waters of Elkhorn Slough, California. Remote Sensing. 2019; 11(14):1664. https://doi.org/10.3390/rs11141664
Chicago/Turabian StyleDierssen, Heidi M., Kelley J. Bostrom, Adam Chlus, Kamille Hammerstrom, David R. Thompson, and Zhongping Lee. 2019. "Pushing the Limits of Seagrass Remote Sensing in the Turbid Waters of Elkhorn Slough, California" Remote Sensing 11, no. 14: 1664. https://doi.org/10.3390/rs11141664